
Type IV pilus is a dynamic structure - PowerPoint PPT Presentation
1 / 60
Title:
Type IV pilus is a dynamic structure
Description:
Type IV pilus is a dynamic structure. PilT mutant have extralong pili that are TM ... GAGs bound to cell surface proteoglycans. Requires Gly-trp repeats of InlB ... – PowerPoint PPT presentation
Number of Views:549
Avg rating:3.0/5.0
Title: Type IV pilus is a dynamic structure
1
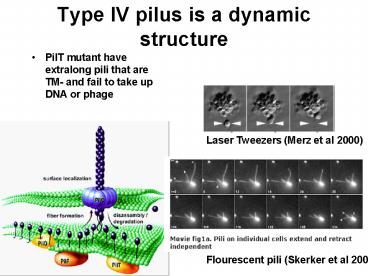
Type IV pilus is a dynamic structure
- PilT mutant have extralong pili that are TM- and
fail to take up DNA or phage
2
Neisseria pathogenesis
- Two human pathogens
- Neisseria gonorrhea STD
- Neisseria meningiditis meningitis, sepsis
- Humans are the only reservoir
- No animal model
3
Diverse, redundant adhesins
- Type IV pili antigenic variation
- CD46 is a receptor
- Opa proteins phase variation (on/off),
antigenic variation - 2 classes of receptor HSPGs, CD66
- Different subclasses of CD66 receptor use
different internalization pathways - Required for human infections
- Opc protein phase variation via mutations in
poly-C tract in promoter region - LOS
- Porins
4
Antigenic variation
PilS genes
Reciprocal recomb Recomb during
transformation Recomb between chromosomes
PilE gene
Phase variation
ATG__________GGGGGGGG____Stop (41
aa) ATG__________GGGGGGGGG_________________stop
(991 aa)
5
Adhesion Invasion
- In vitro systems
- Organ culture (FTOC, NPOC)
- Tissue culture T84 cells
- Initial stages attach as microcolonies
- Elongation of microvilli/filopodia towards
bacteria - Motility dispersal over epithelial surface
- Later times, disperse from microcolonies, pili
disappear, intimate association with host cell
membrane - Engulfment, invasion, transcytosis
6
- Pilus-induced cortical plaques
- Begin within min, lasts for hrs
- PTyr, actin, ezrin, transmembrane glycoproteins
- PilT dependent
7
Pilus-mediated host cell signaling
- Piliated GC/MC or semipurified pili trigger
cysotolic Ca flux. - Requires PilC1
- Blocked by antibodies to CD46
- Depletion of Ca2i ? ?bacterial adherence
- May stimulate lysosomal exocytosis and release of
Lamp1 (substrate for IgA protease) - Recall T. cruzi!
8
Type IV pili induces host signaling
- Host micorarrays compared wt to pilT mutant
- Upregulated MAPK and other stress-activated
pathway and induces genes that protect cells from
apoptosis - Can be mimicked by applying artificial force on
membrane
9
(No Transcript)
10
Two classes of Opa receptors
- HSPGs bind some Opa variants ? binding/internaliza
tion - Localized recruitment of HSPG receptors, F-actin,
P-tyr - Stimulation of Lipid hydrolysis enzymes PC-PLC
and Sphingomyelinase - CD66
- Multiple modes of entry
11
Different CEACAMS specify different entry modes
CEACAM 3 CEACAM 1
12
Neisseria exemplify diversity
- Multiple adhesins/receptors
- Antigenic phase variation
- Multiple pathways of entry
13
Bacterial entry into cells
- Provides access to nutrient-rich environment
- Protection from immune system
- Access to new or deeper tissues
- Must survive in new hostile environment
14
Why bacteria might NOT want to enter into cells
- Harsh environment
- Limited nutrients and iron
- Presentation to immune system
- Limited possibilities for sex or other forms of
genetic exchange
15
Routes of invasion
- Phagocytosis (actin-dependent)
- Entry into professional phagocytes
- IgFc-receptor
- C3b CR1 receptor
- iC3b CR3/CR4 receptor
- Accum of complement components w.o. being lysed
- Delivery to lysosome unless pathogen interferes
with intracellular trafficking - Induced endocytosis/phagocytosis (actin-dep)
- Entry into non-professional phagocyte via
pathogens ligand - Active induction of internalization
16
- Active invasion (actin-independent)
- Active entry without triggering host uptake
pathways - Apicomplexan parasites have motile invasive
stages (zoites) - Probably does not involve specific adhesins
- Route of entry specifies endosomal fate
- Toxoplasma subcellular localiz depends on route
of entry
17
Zipper Hypothesis
Phagocytosis Ingestion of solid material gt 1m
in diameter
Phagocytosis requires repeated sequential
interactions between the phagocyte membrane and
target particle
Implications Cell changes and signaling events
are very local Exquisite cellular discrimination
among surface receptors
18
Multiple stepsMultiple steps at which microbes
can interfere
- Early signaling events
- FcG phosphorylation of ITAM by Src-gtdocking of
SH2 proteins (Syk)-gtPI3K activation, PIP2-gtPIP3,
DAG, PKC - Actin polymerization via Rho GTPases
- Phagosome maturation
- Kiss and run
- Reactive oxygen generation
19
Phagocytosis is complexDictyostelium Genetics
Cardelli, Traffic, 2000
20
Mammalian phagocytosis complex and dynamic
21
Distinct GTPase Effectors in Different Modes of
Phagocytosis
Type III effectors can modulate GTPases and
affect phagocytosis
22
Effector mechanisms resulting from phagocytic
receptor engagement
Fc receptor engagement- proinflammatory CR3-
partially inflammatory Proinflammatory effector
mechanisms NADPH Oxidase (CGD) iNOS PLA2
Recruitment of TLRs TNFa, IL-1, etc.
Recruitment of effector mechanisms can be
blocked by pathogens
23
Entry into non-prof phagocytes
- Bacterial adhesins that mimic eukaryotic
signaling molecules - Yersinia, EPEC, Listeria
- Zipper
- Deliver/inject bacterial molecules that
manipulate host cell cytoskeleton - Salmonella, Shigella, Helicobacter
- trigger mechanism (resemble growth-factor-induce
d membrane ruffling
24
2 modes of entry
Zipper Bacterial ligand binds to host cell
receptor
Trigger Bacterial injects molecules into host
cell
25
Steps in Zipper uptake
- Contact adherence
- Actin independent
- Receptor clustering
- Phagocytic cup formation
- Actin polymerization
- Membrane extension
- Phagocytic cup closure and retraction
- Actin depolymerization
26
Yersinia diseases
- Y. pestis
- Bubonic plague
- Transmitted by flea
- Y. pseudotuberculosis, Y. entercolitica
- Both cause food-borne GI illnesses
- Oral-fecal transmission
- First understanding of bacterial internalization
- Type III secretion discovered inhibits
internalization - Mostly extracellular pathogen
27
- Isberg Falkow 1985
- Cosmid library of Y. pseudotb introduced into E.
coli K12 - Selected for ADHESION
- Screened for INVASION using Gentamycin protection
assay, confirmed by EM - Identified the inv gene
- Yersinia ?inv mutant defective in binding
invasion - Gent protection assay identified another invasin,
Ail - WHY MIGHT THIS APPROACH NOT WORK?
28
Inv gene product
- 103 kD OMP
- Attachment and entry functions cannot be
separated - C-terminal 192 aa is necessary sufficient to
direct entry
Y
MBP-Inv fusion
Anti-MBP
Y
SA
Protein A
29
b1 integrins are the receptor
- Made affinity column of soluble form of inv
- Biotinylated solubilized membrane extracts b1
integrins - Abs to b1 integrins block Yersinia invasion
- Suggests that entry into non-phagocytic cells
related to entry into phagocytic cells (CR are
comprised of integrins) - b1 integrins are receptors for Fn
Y
Y
Y
30
Why does the same receptor bind Fn but
internalize Inv?
- Inv Fn recognize different portions of b1
integrins? - Abs that block Inv binding also block Fn binding
- RGD peptides inhibit both
- Fn is a competitive inhibitor of Inv binding
- Binding and invasion domains of Inv cannot be
genetically separated - Is it just affinity?
- Bacteria coated with anti-a5b1 mAbs uptake
dependent upon affinity of mAbs for receptor
31
Inv Fn have very different structures
- Conserved location of Critical Asp residue
- Convergent evolution
32
- Physical nature of interaction
- High affinity interaction Kd of Fn-a5b1
100-fold greater than for Inv-a5b1 - Invasin monomers multimerize multimerization is
required for uptake - Increased local concentration of integrin
receptors
33
- Lectin domain of Inv is inflexible may stabilize
interaction w/receptor
34
Proof of zipper model
35
Downstream events
- b1 integrins bind talin, a-actinin, FAK,
paxillin, IPCA-1 - b1 integrin cytoplasmic tail req for bacterial
uptake - Some clues provided by type III secreted
anti-invasins - YopH tyrosine phosphatase that targets FAK,
Cas, paxillin, Fyb - YopE Rho-family GTPase activating protein (GAP)
- Cdc42 independent
- Rac activation of Arp 2/3, independent of N-WASP
36
How does Yersinia access b1 integrins?
- M cells are shallow cells specialized in
transcytosis of antigens - Overlay Peyers patches
- Many pathogens access sub-intestinal tissues via
M cells
37
How does Yersinia access b1 integrins?
- B1 integrins expressed on the BL surface of
epithelial cells.
38
Other Yersinia adhesins pH6ag, Ail Yad A
- Also required for binding/or and internalization
- Differential expression
- Inv expressed at RT
- Ail expressed at 37
- Ail mutants show cell-type specific invasion
defect - YadA binds Fn
- pH6 ag binds glycosphingolipids
39
Role of multiple adhesins in vivo
Wt Inv
- InvA mutant is delayed in colonizing PPs (PO)
- LD50 not affected
- YadA mutant dec virulence PO or IP
- InvAil double mutant is avirulent
Intragastric inoculation
40
Yersinia inhibits its own invasion using type III
secreted effectors
- YopE GAP for Rho, Rac, Cdc42
- YopH tyrosine phosphatase, targets focal
adhesion proteins (p130cas, FAK, paxillin, FyB,
SKAP-HOM) - YopT cysteine protease that cleaves isoprenoid
moiety of RhoA at C-terminus - YopO/YpkA serine threonine kinase activated by
G actin and binds to Rho GTPases - Roles in vivo Macrophages? Lymphocytes?
Suppression of oxidative burst? Ca signaling?
41
(No Transcript)
42
Inv homologs are found in other GNs
- Intimin (EPEC, EHEC, Citrobacter freudii)
- Located on LEE PAI
- Homology in N-terminal 500 aa (OM localization
export of C-terminus
Intimin Invasin
43
EPEC
- Common cause of diarrhea in children in
developing countries - Causes attaching/effacing lesions
- Several virulence factors
- Type IV bundle forming pilus (plasmid-encoded)
- Intimin adhesin
- Type III secretion system
- Type III secreted receptor tir
- Other type III secreted proteins
44
EPEC translocates its own receptor (Tir) for
intimin
45
(No Transcript)
46
EPEC inhibits its own invasion
PI3 Kinase
47
EPEC
- EPEC uses intimin as a ligand for binding
- Secretes and translocates its own receptor into
the host cell (Tir) - Causes interesting cytoskeletal rearrangements
- Inhibits its own invasion via type III secreted
proteins - Tir has similarities to H. pylori CagA
- Translocated into host cell membrane
- Variable tyrosine phosphorylation
- Does EPEC move from pedestal to pedestal along
surface of cells? - How does this relate to disease (diarrhea?)
48
(No Transcript)
49
Listeria
- Causes sepsis and meningitis in immunocompromised
adults, abortion in pregnant women, neonatal
infections, rarely GI illness - Food-borne
50
possibly the most famous work of art in
microbial pathogenesis
51
Internalin a family of adhesins
- Screened Tn library of L. monocytogenes for
mutants UNABLE to invade - Defined the Internalin gene family, a family of
23 genes - InlA B are necessary for invasion
- InlA expressed in L. innocua or enterococcus is
SUFFICENT to confer internalization - InlA or InlB is SUFFICENT for beads to be
internalized
GW domains Cell surface Association
52
The real thing
InlA-coated beads
53
E-cadherin is a receptor for Internalin A
- Affinity chromatography using InlA
- Identified E-cadherin
- Ca-dependent cell-cell adhesion through
homophilic interactions between extracellular
domain - BL localization!!!
- Cytoplasmic domain interacts w/catenins actin
cytoskeleton - N-cadherin does not work
54
- LRR and IR regions of InlA sufficient
- Ectodomain of E-cadherin is sufficient for
bacterial adherence - Cytoplasmic domain of E-cadherin is required for
InlA-mediated internalization - Fusion protein of E-cadherin ectodomain fused to
C-terminus of a-catenin sufficient to promote
InlA-mediated entry
55
How can E-cadherin mediate cell-cell contacts and
bacterial entry?
- In cell-cell contact, adjacent cell counteracts
force generated by actin-cadherin complex - For bacteria or InlA-coated beads, no
counter-acting force-gtbacterial entry - Also, multiple E-cadherin-InlA contacts-gt
zippering
56
Using receptor hetergeneity to create a better
animal model
- InlA is not sufficient to mediate internalization
into mouse cells - Chicken L-cam
- hEcadP16
- mEcadE16 -
- hEcadE16E -
- mEcadE16P
- Explains why no role for InlA
- in mouse model of listeria!
57
Transgenic mouse expressing E-cadherinE16P are
permissive for Listeria infection
- Strain Animal LD50 IV LD50 po
- Wt Mouse 105 gt5 x 1010
- ?inlA Mouse 105 gt5 x 1010
- Wt Guinea pig 5 x 107 1011
- ?inlA Guinea pig 5 x 107 gt 1011
- Wt Mouse E16P lt 5 x 1010
- ?inlA Mouse E16P gt5 x 1010
58
InlB-mediated internalization 3 receptors?
- gC1q-R by affinity chromatography
- Requires Gly-trp repeats of InlB
- Subcellular localization function of gC1q-R
controversial - gC1q-R and HSPG binds GW domains and may
dissociate InlB from bacterial surface to host
cell surface - C-Met (HGF receptor) (educated guess based on
activation of specific signaling pathways - PO4 of Shc, Gab1, Cbl
- PI3K and c-Met recruited to site of bacterial
contact - Membrane ruffling sensitive to wortmannin,
genistein, CytD - InlB interacts through LRR
- GAGs bound to cell surface proteoglycans
- Requires Gly-trp repeats of InlB
- May enhance interaction with C-met
59
- InlB necessary sufficient for activation of PI
3-kinase and tyrosine phosphorylation of Gab1,
Cbl, Shc
- Vav (GEF)-Rac-PAK- WAVE-Arp2/3, cofilin -LIM
kinase recruited, then cofilin inactivated by LIM
kinase - HGF InlB bind to different portions of the
C-met receptor
60
Unanswered questions
- What is the relative importance of InlA and InlB
in vivo? - How does Listeria access E-cadherin, a BL protein
- What is the relationship of gC1qR and c-met?
- How do two closely related ligands trigger
different pathways - Are the pathways synergistic?
- E-cadherin and C-met are both at cell junctions
- How does signaling result in internalization?
Recommended
CrystalGraphics Presentations





























