
10.20 Summary of experiments by Nieuwkoop and by Nakamura and Takasaki, showing mesodermal induction by vegetal endoderm (Part 1) - PowerPoint PPT Presentation
1 / 29
Title:
10.20 Summary of experiments by Nieuwkoop and by Nakamura and Takasaki, showing mesodermal induction by vegetal endoderm (Part 1)
Description:
Title: Developmental Biology 8/e Last modified by: KOSLAB Created Date: 1/28/2003 8:12:42 PM Document presentation format: On-screen Show Company – PowerPoint PPT presentation
Number of Views:251
Avg rating:3.0/5.0
Title: 10.20 Summary of experiments by Nieuwkoop and by Nakamura and Takasaki, showing mesodermal induction by vegetal endoderm (Part 1)
1
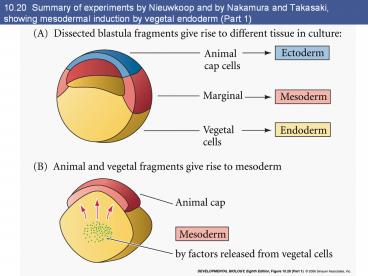
10.20 Summary of experiments by Nieuwkoop and by
Nakamura and Takasaki, showing mesodermal
induction by vegetal endoderm (Part 1)
2
10.20 Summary of experiments by Nieuwkoop and by
Nakamura and Takasaki, showing mesodermal
induction by vegetal endoderm (Part 2)
3
10.21 Experiments on 64-cell amphibian embryos
demonstrated that the vegetal cells underlying
the prospective dorsal blastopore lip region are
responsible for causing the initiation of
gastrulation
4
10.22 The regional specificity of mesoderm
induction can be demonstrated by recombining
blastomeres of 32-cell Xenopus embryos
5
10.23 The role of Wnt pathway proteins in
dorsal-ventral axis specification
- Early 2 cell stage ß-catenin in orange
- Dorsal side of a presumptive blastula and nuclear
ß-catenin - No ß-catenin in the ventral side
- ß-catenin dorsal localization through gastrula
stage
6
10.24 Model of the mechanism by which the
Disheveled protein stabilizes ?-catenin in the
dorsal portion of the amphibian egg (Part 1)
7
10.24 Model of the mechanism by which the
Disheveled protein stabilizes ?-catenin in the
dorsal portion of the amphibian egg (Part 2)
8
10.25 Summary of events hypothesized to bring
about the induction of the organizer in the
dorsal mesoderm
Goosecoid appears to be essential for specifying
the dorsal mesoderm. Goosecoid expression occurs
when there is a synergism between these proteins
and TGF-ß signal secreted by vegetal cells.
9
10.26 Model for mesoderm induction and organizer
formation by the interaction of b-catenin and
TGF-b proteins
Nodal related gene
Ventral and lateral mesoderm
10
Functions of the organizer (page 312)
- While the Nieuwkoop center cells remain
endodermal, the cells of the organizer become the
dorsal mesoderm and migrate underneath the dorsal
ectoderm. - Once the dorsal portion of the embryo is
established, the movement of the involuting
mesoderm establishes the anterior-posterior axis.
- The endomesoderm that migrates first over the
dorsal blastopore lip give rise to the anterior
structures. - The mesoderm migrating over the lateral and
ventral lips forms the posterior structures.
11
- The organizer contribute to four cell types
- Pharyngeal endoderm
- 2) Head mesoderm (prechordal plate)
- 3) Dorsal mesoderm (primarily the notochord)-
induces the hindbrain and trunk - 4) Dorsal blastopore lip- forms the dorsal
mesoderm and eventually becomes the chordaneural
hinge that induces the tip of the tail
Induce the forebrain and midbrain
12
10.27 Ability of goosecoid mRNA to induce a new
axis
The Nieuwkoop center activates the goosecoid gene
in the organizer tissues.
- Gastrula, one blastopore lip
- Gastrula, two blastopore lips,
- goosecoid was injected
C) Goosecoid injected, 2 axes and controls D)
Twinned embryo produced by goosecoid injection
13
10.28 Neural structures induced in presumptive
ectoderm by newt dorsal lip tissue, separated
from the ectoderm by a nucleopore filter with an
average pore diameter of 0.05 mm
- The epidermis that is induced to form, not the
neural tissue. - The ectoderm is induced to become epidermal
tissue by binding bone BMPs. - The nervous system forms from that region of the
ectoderm that is protected from epidermal
induction.
- The default fate of the ectoderm is to become
neural tissue - Certain parts of the embryo induce the ectoderm
to become epidermal tissue by secreting BMPs
14
10.29 Rescue of dorsal structures by Noggin
protein
- Injection of Noggin mRNA into 1-cell, UV-radiated
embryos completely rescues dorsal development. - Noggin induces dorsal ectoderm to form neural
tissue - Noggin dorsalizes mesoderm cells that would
otherwise contribute to the ventral mesoderm. - The development of dorsal structures is a
dosage-dependent.
15
10.30 Localization of noggin mRNA in the
organizer tissue, shown by in situ hybridization
- Noggin mRNA is first localized in the dorsal
blastopore lip region and then becomes expressed
in the notochord. - Noggin binds to BMP4 and BMP2 and inhibits their
binding to receptors.
- At gastrulation at dorsal marginal zone
- When cells involute in the dorsal blastopore lip
- During convergent estension in the precursors of
notochord - Extend beneath the ectoderm in the center of the
embryo
16
10.31 Localization of chordin mRNA
Chordin was found to be localized in the dorsal
blastopore lip and later in the notochord. Of all
organizer genes, chordin is the one most acutely
activated by ß- catenin. Chordin binds to BMP2
and BMP4 and prevents their complexing with their
receptors.
17
Follistatin and BMPs
- - Follistatin is also transcribed in the dorsal
blastopore lip and notochord. - - Follistatin is an inhibitor of both activin and
BMPs, causing ectoderm to become neural tissue. - In Xenopus, the epidermal inducers are BMPs
(BMP4, BMP2, BMP7) and some relatives such as
ADMP (anti-dorsalizing morphogenic protein). - BMP4 induced ectodermal cells to become epidermal.
18
10.32 Model for the action of the organizer
Thus, the epidermis is instructed by BMP
signaling, and the organizer works by blocking
that BMP signal from reaching the ectoderm above
it.
19
10.34 Regional specificity of induction can be
demonstrated by implanting different regions
(color) of the archenteron roof into early
Triturus gastrulae
20
10.35 Regionally specific inducing action of the
dorsal blastopore lip
21
10.36 Paracrine factor antagonists from the
organizer are able to block specific paracrine
factors to distinguish head from tail (Part 1)
22
10.36 Paracrine factor antagonists from the
organizer are able to block specific paracrine
factors to distinguish head from tail (Part 2)
23
10.37 Cerberus mRNA injected into a single D4
blastomere of a 32-cell Xenopus embryo induces
head structures as well as a duplicated heart and
liver
Induction of the most anterior head structures
could be accomplished by a secreted protein
called Cerberus which promotes the formation of
cement glands (the most anterior region of the
tadpole ectoderm), eyes, and olfactory placodes.
Cerberus mRNA injected in a vegetal ventral
Xenopus blastomere originated head structures.
24
10.38 Xwnt8 is capable of ventralizing the
mesoderm and preventing anterior head formation
in the ectoderm
- Frzb and Dickkopf are synthesized in the
involuting endomesoderm (the most anterior
portion of the organizer). - Frzd is a small, soluble form of Frizzled that
can bind to Wnt. - - Dickkopf also appears to interact directly with
the Wnt receptors, preventing Wnt signaling.
25
10.39 Insulin-like growth factors enhance
anterior neural development
IGFs are required for the formation of the
anterior neural tube with its brain and sensory
placodes. IGFs accumulate in the dorsal midline
and are specially prominent in the anterior
neural tube (A).
B) Igf2 injected into ventral marginal zone
blastomeres showing ectopic headlike structure.
C) Control and IGF inhibitor
26
10.40 The Wnt signaling pathway and
posteriorization of the neural tube (Part 1)
The primary protein involved in posteriorizing
the neural tube is thought to be a member of the
Wnt family of paracrine factors, most likely
Xwnt8.
- It appears that a gradient of Wnt proteins is
necessary for specifying the posterior region of
the neural plate. - - In Xenopus, a gradient of Wnt signaling and
ß-catenin is higher in the posterior and absent
in the anterior.
27
10.40 The Wnt signaling pathway and
posteriorization of the neural tube (Part 2)
- There appear to be two major gradients in the
amphibian gastrula - - BMP gradient that specifies the dorsal-ventral
axis and - - Wnt gradient that specify the
anterior-posterior axis
28
10.41 Model of organizer function and axis
specification in the Xenopus gastrula
29
10.42 Pitx2 determines the direction of heart
looping and gut coiling
The embryo has a right-left axis. The crucial
event in this axis formation is the expression of
a nodal gene in the lateral plate mesoderm on the
left side of the embryo. In Xenopus, this gene
is Xnr1. It seems that the microtubules are
involved in the establishment of the Xnr1
expression pattern. Vg1 seems to processed into
its active form predominantly on the left hand
side of the embryo. Vg1- Xnr1- activates
pitx2 Pitx2 persists on the embryos left side as
the head and gut develop, controlling their
respective positions.
Recommended
CrystalGraphics Presentations





























