
Diapositiva 1 PowerPoint PPT Presentation
1 / 27
Title: Diapositiva 1
1
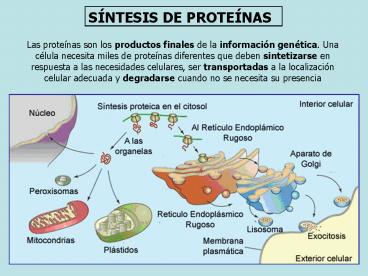
SÍNTESIS DE PROTEÍNAS
Las proteínas son los productos finales de la
información genética. Una célula necesita miles
de proteínas diferentes que deben sintetizarse en
respuesta a las necesidades celulares, ser
transportadas a la localización celular adecuada
y degradarse cuando no se necesita su presencia
2
SÍNTESIS DE PROTEÍNAS Y CÓDIGO GENÉTICO
En qué lugar se sintetizan las proteínas?
Experiencias con aminoácidos radioactivos
determinaron que se realizaba en los ribosomas
Cómo se unen los aminoácidos? Los aminoácidos
son activados (ATP) antes de incorporarse al
polipéptido, uniéndose a un ARNt específico
(complejo aminoacil-ARN) mediante
aminoacil-ARNt-sintetasas específicas
3
SÍNTESIS DE PROTEÍNAS Y CÓDIGO GENÉTICO
4
SÍNTESIS DE PROTEÍNAS Y CÓDIGO GENÉTICO
Cada triplete de bases o codón representa a un
aminoácido o indica el final de la cadena. Los
codones no están separados, sino contiguos, por
lo que la secuencia de una proteína está definida
por una secuencia lineal de codones contiguos.
5
CÓDIGO GENÉTICO Marcos de Lectura
El primer codón de la secuencia establece el
marco de lectura, en el que empieza un nuevo
codón cada tres residuos nucleotídicos. En este
esquema existen tres marcos de lectura posibles
AAUCCGGACUUACGUUAACGGUACAGUAC (1er.
marco) AAUCCGGACUUACGUUAACGGUACAGUAC (2do.
marco) AAUCCGGACUUACGUUAACGGUACAGUAC (3er.
marco)
Una vez establecido el marco de lectura, los
codones se traducen ordenadamente sin
solapamiento ni puntuación hasta que se encuentra
un codón de terminación. Los otros dos marcos de
lectura posibles dentro de un gen normalmente no
contienen información genética útil, pero hay
excepciones. En algunos virus la misma secuencia
produce dos proteínas distintas al utilizar
diferentes marcos de lectura.
6
SÍNTESIS DE PROTEÍNAS Ribosomas y ARNt
7
SÍNTESIS DE PROTEÍNAS Ribosomas y ARNt
Los ARNt tienen entre 73 y 93 residuos
nucleotídicos. Como mínimo cada ARNt tiene ocho
bases modificadas, muchas de las cuales son
derivados metilados de las bases principales
Hay al menos un ARNt para cada aminoácido, pero
para algunos aminoácidos se requiere más de un
ARNt
El bucle (o brazo) de la izquierda reconoce su
lugar en el ribosoma. El de la derecha opera en
el reconocimiento de la aminoacil-ARNt sintetasa
correspondiente. El anticodón reconoce la
ubicación del ARNt en el ARN m.
8
SÍNTESIS DE PROTEÍNAS Fases de la Traducción
La síntesis de polipéptidos empieza siempre en el
extremo amino (NH2) y progresa en el sentido de
la síntesis hacia el extremo carboxilo (CO.OH).
Este patrón se ha confirmado en numerosos
experimentos y es válido para todas las síntesis
en todas las células
9
Etapas de la Síntesis de Proteínas Inicio
La fijación de un factor de inicio a la subunidad
meno impide que las dos subunidades se unan
prematuramente. Luego se fija el ARNm a la
subunidad menor, de modo que el codón de inicio
(AUG, que codifica para formil metionina) se une
en un lugar preciso, porque del lado 5 del ARNm,
a 8-13 pares de bases hay una secuencia que se
unirá con la secuencia complementaria del ARNr de
la subunidad ribosómica menor. Luego de unido el
ARNt que transporta a la fMet se fija la
subunidad ribosómica mayor.
10
Etapas de la Síntesis de Proteínas Elongación
El aminoacil-ARNt que transporta el segundo
aminoácido (Val) se ubica en el sitio A y luego
se forma el primer enlace peptídico entre la fMet
(unida al sitio P) y la Val del sitio A. Esta
reacción produce un dipeptidil-ARNt en el sitio A
y un ARNt descargado en el sitio P. El
siguiente paso es la translocación el ribosoma
se desplaza un codón hacia el extremo 3, que
provoca el traslado del dipeptidil-ARNt al sitio
P, dejando el A libre para la ubicación del
siguiente aminoácido (Phe). Simultáneamente el
ARNt desacilado (el que estaba unido a la fMet)
se desprende del sitio P, volviendo al citosol.
11
Etapas de la Síntesis de Proteínas Terminación
La cuarta fase de la síntesis peptídica está
determinada por la presencia de uno de los tres
codones de terminación del ARNm (UAA, UAG o UGA),
que desencadenan los siguientes procesos
sucesivos (1) la hidrólisis del enlace
peptidil-ARN terminal, (2) la liberación del
polipéptido terminado y (3) la disociación de las
dos subunidades ribosómicas, preparando la
iniciación de un nuevo ciclo de síntesis
polipeptídica.
12
SÍNTESIS DE PROTEÍNAS Esquema de la Traducción
Los ARNt reconocen codones mediante apareamiento
de bases entre el codón del ARNm y una secuencia
de tres bases de ARNt denominada anticodón. Los
dos ARN se aparean de forma antiparalela,
apareándose la primera base del codón (siempre
leída en dirección 5?3) con la tercera base del
anticodón.
13
SÍNTESIS DE PROTEÍNAS Esquema de la Traducción
14
SÍNTESIS DE PROTEÍNAS Eficiencia de la Traducción
Tanto a partir de células procarióticas como
eucarióticas se pueden aislar grandes
agrupamientos de 10 a 100 ribosomas unidos a la
misma molécula de ARNm, denominados polisomas o
polirribosomas, que representan diferentes
estadios de la traducción de la señal contenida
en el ARNm, que es traducida simultáneamente por
muchos ribosomas próximos unos a otros, lo que
permite una utilización muy eficiente del ARNm.
15
Modificaciones Postraduccionales
Modificaciones N- y C-terminales. Todos los
polipéptidos empiezan con f Met (procariotas) o
Met (eucariotas), pero en la mayoría de los casos
son eliminados. Un 50 de las proteínas
eucarióticas tienen el extremo N-terminal
acetilado. Los residuos C-terminales son
modificados con menor frecuencia.
Pérdida de secuencias señal. Péptidos cortos (15
a 30 aminoácidos) que dirigen a la proteína hacia
su destino final. Son eliminados por proteasas
durante la síntesis en el RER
Modificación de amino-ácidos. Los grupos -OH de
Ser, Tre y Tyr pueden ser fosforilados. El
carboxilo de Asp y Glu pueden formar amidas (Asn
y Gln). Lys es frecuentemente metilada o
hidroxilada, como Pro, en la formación del
colágeno.
16
Modificaciones Postraduccionales
.Unión de cadenas laterales glucídicas. La unión
es covalente con residuos Asn (oligosacáridos
unidos por enlaces N), como en el caso de las
proteínas que entran el RER (a la derecha) o
residuos de Tre o Ser (oligosacáridos unidos por
enlaces O) como en los proteoglicanos de la
matriz extracelular (abajo).
17
Modificaciones Postraduccionales
Adición de grupos isoprenilo. Los grupos
isoprenilo provienen de la biosíntesis del
colesterol. Las laminas (proteínas de la lámina
nuclear) y muchas proteínas de membrana están
preniladas.
Adición de grupos prostéticos. Un ejemplo es el
grupo hemo del citocromo c (arriba) y de la
hemoglobina (izquierda).
18
Modificaciones Postraduccionales
Modificación proteolítica. La insulina, las
proteasas pepsina, tripsina y quimotripsina y el
colágeno se sintetizan como precursores inactivos
(proinsulina, pepsinógeno, tripsinógeno,
quimotripsinógeno y protocolágeno,
respectivamente) que luego deben ser hidrolizados
parcialmente para convertirse en productos
biológicamente activos. Formación de puentes
disulfuro. Pueden ser intra- o intercatenarias y
se supone que ayudan a las proteínas que deben
ser exportadas a mantener su estructura nativa.
19
SISTEMA DE MEMBRANAS INTERNAS
Lla célula de los eucariotes posee un laberinto
de membranas internas que rodean al núcleo y que
se extienden a distintas partes del citoplasma.
En algunas células ocupa gran parte del volumen
citoplasmático y constan de una serie de láminas
estrechamente empacadas y plegadas en forma
aplanada, de manera que dan origen a diversos
compartimientos dentro del citoplasma. Las
cavidades formadas por las láminas de las
membranas se denominan cisternas
20
Retículo Endoplásmico (RE)
Hay dos tipos de RE el RE rugoso (RER), que
tiene ribosomas en su superficie y por lo tanto
en las microfotografías electrónicas ostenta un
aspecto rugoso, y el RE liso (REL), que carece de
ribosomas, por lo que la superficie externa de su
membrana tiene una apariencia lisa. Las membranas
del REL tienden a formar túbulos mientras que
las membranas del RER forman en general sacos
aplanados. No son orgánulos independientes por
microscopía electrónica se demuestra una
continuidad entre el lumen de ambos.
21
Retículo Endoplásmico liso (REL)
Es el principal sitio de metabolismo de los
lípidos. Realiza también una función importante
al localizar enzimas desintoxicantes que degradan
sustancias químicas tóxicas o carcinogénicas y
las conviertan en moléculas solubles fácilmente
excretables por el organismo. Algunas líneas
celulares, como las células del hígado, contienen
grandes cantidades de REL. En otras células del
cuerpo el REL llega a ser un componente menor. El
REL de los hepatocitos está también involucrado
en la hidrólisis enzimática de glucógeno, que se
almacena en gránulos asociados al REL
22
Retículo Endoplásmico Rugoso (RER)
El RER juega un papel central en la síntesis de
las proteínas de sus propias membranas y en la
síntesis de proteínas de otras membranas. Muchas
de las proteínas que la célula exporta (como las
enzimas digestivas o las hormonas proteicas) o
aquellas destinadas a otros organelos o a la
propia membrana plasmática se forman en los
ribosomas unidos a la membrana del RER
23
Aparato de Golgi
Muchas de las proteínas secretadas por las
células, así como las que se integran a la
membrana plasmática y las que son dirigidas a
otros organelos del sistema endomembranoso, pasan
a través del aparato de Golgi. Después que estas
proteínas son sintetizadas por ribosomas unidos
al RER se transportan al aparato de Golgi
mediante vesículas de transporte formadas en la
membrana del RER
24
Lisosomas
Pequeños sacos que contienen enzimas digestivas
que degradan lípidos, proteínas, carbohidratos y
ácidos nucleicos, originados dentro y fuera de la
célula. Contienen cerca de 40 enzimas diferentes,
todas hidrolasas activas a pH 5 (proteasas,
lipasas, fosfolipasas, glicosidasas, fosfatasas,
sulfatasas, nucleasas). Se sintetizan en el RER y
son luego modificadas en el aparato de Golgi.
25
Proteasomas
La mayoría de las proteínas que se degradan en el
citosol de las células eucarióticas lo hacen a
través de grandes complejos de enzimas
proteolíticas denominados proteasomas.
26
Vacuolas vegetales
La mayor parte del volumen de muchas células
vegetales están ocupadas por una vacuola central
grande. Participa en el equilibrio de agua,
sirve como acúmulo de desechos que no pueden
excretarse. Algunas células almacenan en sus
vacuolas sustancias tóxicas, y ello disuade a los
animales que mastican las hojas. También pueden
almacenar nutrientes o pigmentos, como en los
pétalos de las flores.
27
Microcuerpos
Son organelos delimitados por una membrana, pero
no derivan del RE y por lo tanto no forman parte
del sistema de endomembranas . En la degradación
de lípidos se produce peróxido de hidrógeno
(H2O2), que es degradado en los peroxisomas,
donde la catalasa utiliza el H2O2 para oxidar
otros sustratos. Los peroxisomas de las células
de hígado y riñón tienen gran importancia en la
desintoxicación de algunos compuestos como etanol.
Las células vegetales contienen un tipo de
peroxisoma que participa en la fotorrespiración y
el otro tipo de microcuerpo es el llamado
glioxisoma, que contiene enzimas que convierten
en azúcares los lípidos almacenados en la
semillas, durante la germinación.
Recommended
