
Bioenergetics PowerPoint PPT Presentation
1 / 36
Title: Bioenergetics
1
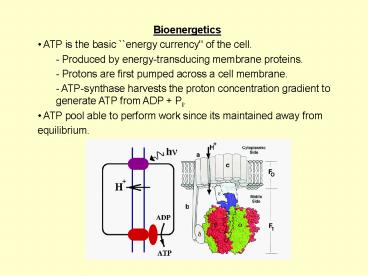
- Bioenergetics
- ATP is the basic energy currency'' of the
cell. - - Produced by energy-transducing membrane
proteins. - Protons are first pumped across a cell membrane.
- ATP-synthase harvests the proton concentration
gradient to generate ATP from ADP Pi. - ATP pool able to perform work since its
maintained away from - equilibrium.
2
- Enthalpy
- The first law of thermodynamics says total
energy conserved - D U q - w
- ie. the change in energy of a system is the heat
absorbed by the system from the surroundings, q,
minus the work done by the system on the
surroundings, w. - Enthalpy defined as
- H U PV
- Convenient for biology since, under constant
pressure (DP 0), - D H is measured as the heat produced or
absorbed - D H D U P D V q - w P D V q
- since at constant pressure w P D V.
3
- Entropy
- Thermodynamics describes the bulk behavior of
reversible systems. - - Key concepts enthalphy Gibbs free energy
entropy. - Entropy
- S kB ln W
(1) - where kB is Boltzman's constant and W is the
number of ways of arranging a system. - Systems adopt their most probably arrangement.
- Corresponds to those with maximum entropy.
- Example Ten gas molecules in a bulb with a
barrier - One way of having all ten in left bulb.
- Ten ways of having nine in left and one in
right. - And n!/l!(n-l)! 10!/(5! 5!) 252 ways
of having five in left and five in right.
4
- Gibbs Free Energy
- Gibbs free energy defined as
- G H - TS
- T is the temperature, and S the entropy.
- For constant temperature and pressure (D T 0,
D P 0) - D G D H T D S
- D H is the changes in enthalpy (ie. heat
exchanged). - Closed system implies constant temperature and
pressure. - Typical experimental conditions.
- Reactions are spontaneous if D G lt 0.
5
Reversible reactions The reaction ATP ? ADP
Pi can be considered as two simultaneous
reactions The consumption of ATP ATP ? ADP
Pi with a rate constant k1 - d/dt
ATP d/dt ADP k1 ATP
(2) in competition with the production of
ATP ADP Pi ? ATP with a rate constant k2
d/dt ATP k2
ADP Pi (3)
6
- When at equilibrium (from Eqs. (2) and (3))
- d/dt ATP k2 ADP Pi - k1 ATP 0
- ie. rate of ATP production rate of ATP
consumption hence - ADP Pi / ATP k1/k2 KATP
hydrolysis (4) - which defines the equilibrium constant for ATP
hydrolysis. - The experimentally measured value is
- KATP hydrolysis 105 M
- In the presence of 10-2 M Pi and 10 -3 M ADP
(approximate values for the mitochondrial
cytoplasm) - ATP 1/K ATP hydrolysis ADP Pi
- 1/105 10-3 10-2 M 10-10 M
- only one molecule of ATP per ten million of ADP
if allowed to run down to equilibrium.
7
- Gibbs free energy in solution
- For the reaction
- A ? B C
- define the mass-action ratio
- G B C / A
(5) - such that G Kequil at equilibrium.
- From thermodynamics (ie. without proof), the
change in Gibbs free energy for a reaction in
solution is - D G - 2.3 R
T log10 Kequil / G (6) - D G0 2.3 R T log10 G
- R the gas constant (R 8.3 J/(mol K)).
- T temperature in Kelvin (K).
- G0' - 2.3 R T log10 Kequil and ' indicates pH 7
(bio. convention). - At equilibrium G Kequil so DG 0.
- A half-proof'' of how Eq. (6) is derived from
Eq. (1) is given as an appendix to Chapter 3 of
Biochemistry, Voet and Voet.
8
- Example ATP hydrolysis reaction
- Use Eq. (5) to determine D G as G is displaced
from equilibrium. - G ADP Pi / ATP
- and
- KATP hydrolysis 105 M
- Suppose in the cell Pi 10-2 M and is (more or
less) constant.
9
- In healthy cells ATP 10-2 M and ADP 10-5
M, such that - G 10-5 10-2 / 10-2 10-5 10-10 KATP
hydrolysis - Hence
- D G -2.3 R T log10 KATP hydrolysis / G
- -2.3 8.3 J/(mol K) 298 K 10 - 57 kJ/mol
- The reaction is so displaced from equilibrium
that in consuming one mol of ATP 57 kJ of work
can be performed.
10
- The standard free energy (25oC pH 7) of
phosphate hydrolysis of ATP to ADP is DG0 '
-30.5 kJ/mol. - Free energy of a pure elements in their standard
state (25oC, 1 atm) and most stable form (eg. O2)
is defined as zero. - Free energy of formation of another substance is
the change in free energy accompanying the
formation of one mol of that substance under
standard conditions. - - DG 0 ' is the difference between the free
energies of formation of ATP versus ATD and Pi
under '' standard'' conditions. - Since 57 kJ/mol gt 30.5 kJ/mol is there a
contradiction? - - It is the displacement of G from equilibrium
which gives the cell its ability to do work! - When ATP is displaced 10 orders of magnitude
from equilibrium it can do more work than that
associated with phosphate bond hydrolysis under
standard conditions. - Entropy the driving force for gas molecules to be
in both bulbs - An entropy advantage of having a mixture of ATP,
ADP, AMP etc. ie. more disordered'' if not a
pure species.
11
- Compare with
- DG0 2.3 R T log10 Kequil
- 2.3 8.3 J/(mol K) 298 K 5
- 28 kJ/mol.
- Difference between 30.5 kJ/mol and 28 kJ/mol is
round-off error (eg. Kequil accurate to only one
decimal place).
12
- Measuring the Gibbs free energy across a membrane
- The Gibbs free energy across a membrane is given
by - D G - F Dy - 2.3 RT log10 (HP /
HN) (7) - - F is Faraday's constant 0.097 kJ/(mol mV).
- - Dy electric potential difference either side
of the membrane. - - HP proton concentration on the positive
side. - - HN proton concentration on the negative
side. - The first term is the energy of the stored
electrostatic potential (ie. like a capacitor). - The second term Gibbs free energy for the
reaction HP ? HN - ie. the pH gradient is a displacement from
equilibrium (Eq. 5).
13
- Measuring D pH
- A weak acid obeys
- HA ? H A-
- with Equilibrium constant
- Ka H A- / HA (8)
- Since HA is neutral it will freely permeate the
membrane. - - Either side of the membrane HAN HAP
- We know HP ? HN due to D pH.
- - Therefore A-P ? A-N at equilibrium.
Note for a weak acid HA gtgt H, A-, so
picture is just illustrative, don't take
literally!
14
- Measuring DpH done by
- Growing organelle's with physiological pH
gradient (A). - Adding A- to the incubation medium (B).
- Stop respiration (eg. rapidly separate organelle
from incubation medium). - Monitoring the fall of A- outside the organelle
(C to D) since HA will diffuse through the
organelle's membrane. - Alternatively use a spectral indicator for A-
as it accumulates inside the organelle (D). - Amount of HA found inside the organelle closely
related to - (almost matching) the excess of H outside the
organelle when - A- added.
15
(No Transcript)
16
- Measuring D y
- Measure the electrostatic potential difference
by having an ion (eg. X) which can freely
permeate the membrane - - Requires a rapid transporter of X.
- Allow X to equilibrate either side of the
membrane as the organelle functions. - - Dy will drive a concentration gradient for X.
- At equilibrium
- DG - F Dy - 2.3 RT log10
(XP / XN) 0. (9)
17
- Solving Eq. (8) for Dy gives
- Dy - 2.3 RT / F log10 (XP /
XN) (10) - Measuring the fall in X outside the
organelle, or increase inside, gives Dy directly.
- Must know internal volume of organelle.
- Must have only one rapid transport mechanism of
X. - X should not disturb pH gradient (ie. X must
be low). - Need a good indicator for X.
18
- First measurements of transmembrane Gibbs free
energy - Mitchell and Moyle (1969) employed pH and K
specific electrodes in anaerobic, low K
incubation - Valinomycin used to transport K.
- Dy determined by K uptake during an O2 pulse.
- Used mitochondrial respiratory chain.
- DpH estimated by simultaneous proton extrusion.
- A value of 228 mV for Dy.
- - Equates to 22 kJ/mol.
- In the mitochondria DpH only makes a small
contribution to total Gibbs free energy. - - Buffer effects protect enzymes from pH
extremes. - In chloroplasts DpH makes a larger contribution.
- - Have very pH robust enzymes in these
organelles.
19
- Does it Add Up?
- First success of the theory was it explained why
charge transporters - (eg. valinomycin, which transports K) kill
ATP-synthesis. - Mitchell and Moyle found Dy 228 mV in
mitochondria. - - Equates to 22 kJ/mol.
- Know the mitochondria maintains the ATP ? ADP
Pi reaction ten orders of magnitude from
equilibrium. - - Equates to 57 kJ/mol energy storage by ATP.
- Believed that 3 to 4 H are pumped in the
mitochondria per ATP molecule synthesized. - - Between 66 and 88 kJ/mol of energy available
for each 57kJ/mol ATP molecule generated. - - Surprisingly efficient!
- This finding crucial, since if DG did not add
up the whole theory would need to be abandoned.
20
- Proton pumping
- Established that energy transduction proceeds by
proton pumping. - Proton motive potential then harvested by
ATPsynthase. - How can it possibly work in detail?
- - Demands structural information.
21
- Classical proton pump
- A binding site for a proton.
- Resting state has very high proton affinity.
- A proton taken up.
- Upon the input of energy
- Conformational change.
- Switches accessibility to the other side of the
membrane. - Dramatically reduces proton affinity of binding
site. - A proton is released.
- Net translocation of a proton.
- - Basic model Jardetzky, Nature 211, 969-970
(1966).
22
- Redox coupled proton pump
- Contain two redox chemistry sites.
- - One either side of the membrane.
- Oxidation of substrate S1
- Reduces the enzyme.
- Electrons flow to second redox site.
- Protons released to one side of membrane.
- Reduction of substrate S2
- - Protons taken up from other side of the
membrane. - Net translocation of a proton.
- - Basic model Mitchell, Nature 191, 144-148
(1961).
23
- Electron proton movements
- Electrons (or protons) sit within potential
energy wells. - Have specific quantum mechanical''
wavefunctions. - Energy of wavefunction hn (n oscillation
frequency). - Neighboring potential energy wells always
present. - eg. an electron may move to a neighboring
cofactor. - eg. a proton may move to a neighboring
residue/water molecule. - To be probable that the electron/proton moves
- For a given set of nuclear coordinates the total
potential energy of reactants and products must
match. - Efficient quantum-mechanical wavefunction
coupling If hnreactants hnproducts
24
- Rate constants of transfer
- Rates for electron (proton) transfer depend on
- centre-to-centre distance of reactants and
products. - As distance increases energy barrier goes up!
- Probability of fluctuations providing this
energy goes down. - Transfer probability of crossing from one
surface to another depends on extent of
wavefunction overlap. - - eg. Certain angles favored for H-bonds.
- - eg. Electron wavefunctions have structure and
therefore - overlap better at favored orientations.
- - Proteins carefully arranges cofactor positions
and orientations.
25
- Tunneling
- May expect that proteins provide pathways
between electron donors and acceptors. - - In practice not so.
- Electrons pass from one centre to another by
tunneling. - There is a finite probability that the electron
is already in the product's potential well. - - Diagnostic for tunneling is insensitivity to
temperature changes.
26
- Proton Exchange
- Have hydrogen bond partners.
- - If distance and angles favorable have very
rapid exchange of a proton. - - Imagine the forward and reverse reaction
happening continuously. - H-bond networks of charged side chains and water
molecules serve as proton wires''.
27
- What determines where protons lie?
- Define the pKa of a residue/cofactor/water HA
as - pKa - log10 (HA- / HA)
- where the reaction is HA ? H A-.
- Low pKa means relatively high H.
- High pKa means relatively low H (ie.
relatively high OH-). - A number of residues have a pKa in the region
where their - protonation states can change
- - Aspartate (pKa 3.9), Glutamate (pKa 4.1),
Histidine (pKa 6.0), Lysine (pKa 10.5),
Arginine (pKa 12.5), Tyrosine (pKa 10.5),
Cysteine (pKa 10.7). - If two residues in H-bond contact (or linked via
water molecules) the proton will lie on that with
highest pKa. - Several factors perturb the pKa of residues
- Number of H-bonds.
- Distribution of charges (eg. nearby positively
charges can stablilse a negative charge on a
residue and vice versa). - How hydrophobic/hydrophilic the local
environment is.
28
- Classical proton pump
- Resting state has a high affinity proton binding
site. - - H-bonds/local charges etc. stabilise its high
pKa. - Conformational change.
- Switches accessibility to the other side of the
membrane (eg. opens and closes entry/exit
channels). - pKa of proton binding site drops significantly.
- - Loss of H-bonds, movements of charges etc.
- A proton is released.
29
- Flow of electrons
- Similar structural/energy barrier considerations
to proton transfer. - Electron flow faster since e- 1000 times lighter
than H. - Can also have significant tunneling'' through
barriers. - - It is the Oxidation-Reduction (Redox) potential
which is relevant. - General reaction is
- Oxidised ne- m H ? Reduced
- An Oxidised/Reduced pair is called a redox
couple. - In the case of a metal and its salt
- Two half cells of metal electrode and its salt
versus a standard. - Link them by a salt bridge and measure the
potential difference.
30
- The metal electrode which gives up electrons has
the lower electro-potential. - Standard electro potentials defined against a
H2/Pt electrode _at_ 1Mol concentration H (ie. pH
0) and 25oC. - Normal biological convention defines
electropotentials _at_ pH7. - Mid-point potential is that when Oxidised
Reduced - The actual redox potential at pH x is
- Eh,pHx Em, pH x ((2.3 R T)/(n F))
log10(ox / red) - Where Em, pH x is the mid-point potential at pH
x. - Note the energy term arising as Oxidised and
Reduced - change, related to earlier expressions!
31
Some mid-point potentials _at_ pH 7 Oxidised ne-
m H ? Reduced
32
- Common redox groups in biology
- Most enzymes use relatively few redox cofactors.
- Cu centres.
- Single Cu dinuclear (two Cu) centres binuclear
(one Cu, one Fe) centres. - Fe/S complexes.
- - Two Fe2 and two S2-.
- Heme.
- - Normally Fe2 within a porphyrin ring.
- Chlorophyll.
- - Like heme but with Mg2 rather than Fe2.
- Pheophytin.
- - Like heme but with 2H rather than Fe2.
- Quinone/Quinol.
- Redox potentials modified by subtle structural
effects.
33
- Redox coupled proton pump
- Contain two redox chemistry sites.
- Oxidation of substrate S1
- - Has a relatively negative electro-potential.
- - Electrons flow to second redox site.
- Reduction of substrate S2
- - Has a relatively positive electro-potential.
- Net translocation of a proton.
34
- Structural methods
- To understand these processes need structural
information. - Nuclear Magnetic Resonance
- - Has a relatively low upper size limit.
- Electron microscopy.
- - Quite successful for membrane proteins but low
resolution. - Electron microscopy structure of Ca2-ATPase.
- X-ray diffraction.
- - The most successful and high resolution.
- - Difficult to get crystals.
35
- The X-ray approach
- Grow crystals.
- Travel to synchrotrons.
- Collect X-ray diffraction data.
- Interpret electron density.
36
- Summary Lecture 2
- Thermodynamics determines Gibbs free energy of
reactions. - - A reaction spontaneous if DG is negative.
- Measurements of proton gradients consistent with
the energy content of ATP within the cell. - Two types of proton pump
- - For classical proton pumps must consider pKa
values. - - For redox coupled pumps electropotentials the
central idea. - X-ray diffraction the most promising method for
elucidating structural details.
Recommended
