
living things are - PowerPoint PPT Presentation
1 / 41
Title:
living things are
Description:
electrochemical gradient (potential) depends on both ... Iron-sulfur proteins as electron carriers Fe is not ... All participate only in one-electron transfers ... – PowerPoint PPT presentation
Number of Views:54
Avg rating:3.0/5.0
Title: living things are
1
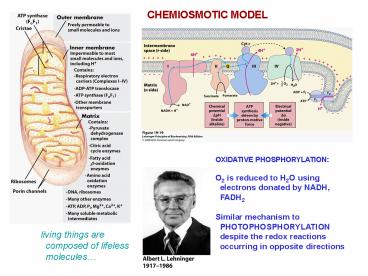
CHEMIOSMOTIC MODEL
OXIDATIVE PHOSPHORYLATION O2 is reduced to H2O
using electrons donated by NADH,
FADH2 Similar mechanism to PHOTOPHOSPHORYLATIO
N despite the redox reactions occurring in
opposite directions
living things are composed of lifeless
molecules
2
Oxidative phosphorylation AND
photophosphorylation -electron transport through
carriers -uphill transport of protons through
a membrane, creating a transmembrane
electrochemical potential -ATP synthesis
driven by this potential ATP synthase
enzyme ________________________________ -electron
transport and ATP synthesis occur in the inner
mitochondrial membrane, adjacent to the
matrix -ADP and Pi are transported into the
matrix from the cytosol, while ATP is
transported out
3
MECHANISMS OF MEMBRANE TRANSPORT
4
Movement of electrically charged solutes across a
membrane electrochemical gradient
(potential) depends on both concentration
difference and membrane electrical potential
5
-Both NAD and NADP are soluble and diffusible in
cells -Substrate is oxidized (two hydrogen atoms
are removed) -2 electrons and one proton are
transferred to NAD/NADP as a hydride
ion -Remaining proton is released to solvent
6
Flavin cofactors can undergo either one or two
electron transfers greater diversity of
reactions than for NAD/NADP Flavin cofactor is
very tightly bound, may be classed also as a
prosthetic group. Non- diffusible in
cells Great variability in reduction potential
depending on which enzyme bound to
7
Ubiquinone (coenzyme Q) lipid-soluble
benzoquinone isoprenoid side-chain for
lipid solubility Ubiquinone has three oxidation
states Like FAD, can function as both a one
and 2-electron carrier Freely diffusible in the
membrane Carries both electrons and protons
(coupling electron flow to proton gradient)
8
Cytochrome c absorption in oxidized and reduced
forms
- Cytochromes as electron carriers these proteins
have Fe-containing - heme prosthetic groups with characteristic
absorption spectra - Sometimes named for their precise absorption
maxima eg, cytochrome b562 - Note covalent attachment of heme in cytochrome c
noncovalent in cytochromes a, b - Fe atom interacts with protein, reduction
potential depends on detailed environment - May be integral membrane proteins, or
membrane-associated
9
4Fe-4S center
2Fe-2S center in ferredoxin
Iron-sulfur proteins as electron carriers Fe is
not associated with heme, but is linked to
inorganic sulfur or enzyme Cys side-chains All
participate only in one-electron transfers 8 or
more Fe-S proteins in the electron transport
chain in mitochondria widely varying reduction
potentials
2Fe-2S center
10
- Standard reduction potentials of all individual
electron carriers - have been determined
- Order of transfer NADH?FADH2?Q?Fe-S?Cytochrome?O2
- Standard order and actual order of transfer may
differ according to - the concentrations of oxidized and reduced
forms in vivo
11
The electron chain functions in four
membrane-bound supramolecular complexes Complexes
can be fractionated and studied individually
12
Electron flow through the four functional
complexes in the mitochondrial inner membrane
13
(No Transcript)
14
ATP synthesis is driven by the favorable downhill
flow of protons from the intermembrane space
back into the matrix
15
Triacylglycerol? glycerol? glycerol-3-P (cytosol)
Path of electron transfer through the first
two complexes Both complexes I and II contain
Fe-S proteins as intermediate carriers Complex
I receives electrons from NADH complex II
receives electrons from succinate Path of
electron transfer from fatty acyl-CoA through
acyl-CoA dehydrogenase, converges at coenzyme
Q
b-oxidation
Note connections to lipid metabolism
16
NADHubiquinone oxidoreductase a proton pump
driven by the energy of electron transfer
(42 polypeptides, 1 flavoprotein, gt6 Fe-S centers)
QH2 is freely diffusible in the bilayer and
goes next to complex III
structure of matrix domain is solved
- NADH H Q ? NAD QH2
- Vectorial transfer of 4 protons matrix to
intermembrane space - Net NADH 5HN Q ? NAD QH2 4HP
These steps must be coupled as the proton
transfer is disfavored and is driven by the
favorable redox energy of the electron transfer-
as a hydride ion from NADH to Q
17
Complex II is the same as succinate dehydrogenase
of TCA cycle Electron flow travels from
succinate to FAD, through 3 Fe-S centers, and
ultimately to Q QH2 then diffuses off to
complex III Four enzyme chains, full structure
known including locations of cofactors Total
pathlength of electron transfer is 40 Å, but
longest individual step is 11 Å
(succinate)
18
- CoQ is diffusible and transports all electrons to
complex III - Electron transport from complex III to complex IV
goes by way of - cytochrome c, which diffuses through the
intermembrane space
19
Complex III Cytochrome bc1 complex
Dimeric complex each monomer has 11
subunits Of the 11, three comprise the core
cytochrome b, cytochrome c1 and Rieske
iron-sulfur protein Dimer interacts with
cytochrome c in the intermembrane space Two
quinone binding sites QP and QN Each is bound
between the monomers Recall that QH2 arrives at
complex III from complexes I and II and now
donates its electrons to cytochromes via a Q
cycle
20
Q Cycle on Complex III
Cytochrome c will move off to complex IV after
it is reduced
1. Two molecules of QH2 are oxidized to Q on the
intermembrane (P) side, releasing a total
of four protons into the intermembrane space. 2.
Each of the two QH2 transfer one electron to
cytochrome c1 via Fe-S, and one electron
to Q via cytochrome b. Two 1-electron transfers
to Q plus uptake of two protons from the
matrix regenerate a QH2 molecule ? net vectorial
proton transport
21
Complex IV Cytochrome oxidase
Binuclear copper center (CuA) two Cu and 2
Cys, resembles 2Fe-2S center
Fe-Cu center contains Fe in two hemes heme a
and heme a3, together with one Cu (CuB). Heme
a3/CuB also form a binuclear center
Reduced cytochrome c diffuses freely from complex
III Electron transfer is from cytochrome c ? CuA
center ? heme a ? heme a3/CuB ? O2 Four
electrons, one at a time, are needed to reduce
molecular oxygen to 2 waters Four protons are
taken up into water and four more are pumped out
of the matrix for every four electrons that
pass through from cytochrome c Net reaction 4
Cyt c (Fe2) 8 HN O2 ? 4 Cyt c (Fe3) 4 HP
2H2O
22
Summary of electron and proton flow
23
Proton gradient across the inner mitochondrial
membrane generates a proton motive force due
to both concentration difference and
electrical potential energy
24
(No Transcript)
25
CHEMIOSMOTIC MODEL
ATP synthesis is driven by the favorable downhill
flow of protons from the intermembrane space
back into the matrix.
26
(ii)
(iii)
(i)
Isolated mitochondria suspended in buffer,
addition of substrate or of ADP/Pi (i)
cyanide blocks electron transport (blocks
cytochrome oxidase function) ?inhibiting
electron transfer to O2 blocks ATP synthesis
(ii) venturicidin is a toxic antibiotic that
inhibits ATP synthase ?inhibiting ATP
synthesis blocks the electron transfer pathways
(iii) dinitrophenol (DNP) is an uncoupler
?allowing protons back into the matrix by a
different mechanism enables continued
oxidation of succinate
ATP synthesis and oxygen consumption depend on
the presence of an oxidizable
substrate Oxidation of succinate requires ATP
synthesis Therefore the two processes are
obligatorily coupled If no ATP synthase, proton
gradient builds until further pumping impossible
27
- DNP and FCCP each have
- a dissociable proton
- They carry protons across
- the inner mitochondrial
- membrane and cause
- dissipation of the gradient.
- Then electron transfer from
- NADH to oxygen occurs,
- but it is not harnessed for
- ATP synthesis anymore
28
- Structure of ATP synthase.
- F1 is a peripheral membrane protein
- and Fo is an integral protein
- Extraction of F1 from the intact
- enzyme still allows electron transport,
- but no proton transport because
- protons immediately leak back in
- Purified F1 added in trans reconstitutes
- F1 alone catalyzes ATP hydrolysis
F1
MATRIX
Fo
INTERMEMBRANE SPACE
Mimic of transition state for ATP synthase ADP
b-oxygen attacks phosphorus of Pi, water is
the leaving group pentacovalent transition
state needs Mg2
29
- ATP synthase binds ATP very tightly 107-fold
tighter than ADP - The greater binding energy equalizes the free
energy of ATP - synthesis with that of ATP hydrolysis when on
the enzyme surface - The proton motive force is necessary not for ATP
synthesis but to - dislodge the formed ATP off the enzyme
surface - The enzyme must cycle between two conformational
states one that - binds ATP tightly and one that allows ATP
release
30
F1
Fo
- Structure of F1 portion of the ATP synthase
- Stoichiometry is a3b3gde, but d and e not seen
- Structures of the b subunits are each slightly
- different because of asymmetric interactions
- with the g subunit.
- One b subunit binds ATP, one binds ADP,
- and one remains empty
31
Structure of the Fo transmembrane
protein Stoichiometry is ab2c10 c subunits are 8
kD, each 2 helices, arranged in two concentric
rings g subunit of F1 points through the center
of the Fo ring In schematic (upper left),
positions and structures of a, b2 and e are
models only
32
F1
- Binding-change mechanism for ATP synthesis
- rotational catalysis
- The three b-subunits of F1 alternate in ATP
synthesis - At any time, one b-subunit binds ATP,
- one binds ADP, and one is empty
- Proton motive force drives rotation of the
central - cylinder (the Fo c subunits) and shaft (the F1
- g and e subunits)
- The g subunit (green) contacts each ab-subunit
- pair as it turns, causing coupled
conformational - changes
- Contact of g with a b-subunit forces it to become
- empty to release its ATP the difficult
step (top) - After ATP is released, 120 rotation causes the
- ADP-bound form to adopt the conformation in
- which ATP is tightly bound, allowing
equilibration - of ATP with ADP on the enzyme (ATP synthesis)
- 360 rotation causes each b-subunit to transit
through
Fo
33
Use of biotin-avidin to watch rotation of ATP
synthase -Fluorescent actin covalently attached
to avidin -Biotin covalently attached to Fo c
subunit -Addition of ATP, or of ADP Pi,
causes rotation in opposite directions -Fluorescen
ce micrographs at 133 msec intervals
discrete jumps are made
34
- ATP synthase is in proximity to 2
- other proteins in the inner mito.
- membrane an antiporter and a
- symporter
- Adenine nucleotide translocase
- transports ADP into matrix and
- ATP into intermembrane space
- favored by proton gradient
- Phosphate translocase, favored
- by electron-transport-generated
- proton gradient, and dissipates it
- by a small amount
35
Oxidation of one NADH in the matrix transports 10
protons Oxidation of one succinate transports 6
protons
36
2 NADH produced in glycolysis could yield either
3 or 5 ATP ? P/O 1.5 or P/O 2.5
37
NADH regenerated
NADH from glycolysis
MALATE- ASPARTATE SHUTTLE
- Complex I accepts electrons from NADH only in the
matrix - This shuttling cycle moves electrons into the
mito. in the form of malate - The intermembrane space is freely diffusible into
the cytosol
38
GLYCEROL-3-PHOSPHATE SHUTTLE
DHAP is reduced to glycerol-3-phosphate in the
intermembrane space, oxidizing NADH Reduced
glycerol-3-phosphate is re-oxidized, reducing FAD
in turn. FADH2 then is bound by a dehydrogenase
in the outer face of the inner membrane Transfers
electrons directly to complex III so generates
only 6 protons per NADH
39
REGULATION OF OXIDATIVE PHOSPHORYLATION
Red/white protein bridging the F1-ATPase is
IF1 IF1 binds to the ADP conformation, freezes
rotation, and blocks ATP hydrolysis under
conditions when there is no oxygen/no electron
transport (heart attack or ischemia) The IF1
is a dimer stabilized only at low pH Anaerobic
cells go to low pH due to lactate
accumulation IF1 inhibition then prevents
wasteful ATP hydrolysis resumption of aerobic
metabolism raises pH, causing IF1 dimer to fall
apart and ATP synthase
40
Regulation of oxidative phosphorylation is also
at the level of ADP concentration responds
to energy charge and is linked in this sense to
glycolysis and to the TCA cycle
41
Uncoupling protein thermogenin allows protons
back into mitochondrial matrix Proton gradient is
dissipated as heat This is desirable in newborn
infants, to generate heat in adipose
tissue Maintains body temperature in newborns
Recommended
CrystalGraphics Presentations





























