
A fundamental principle of development is epigenesis - PowerPoint PPT Presentation
1 / 81
Title: A fundamental principle of development is epigenesis
1
A fundamental principle of development is
epigenesis
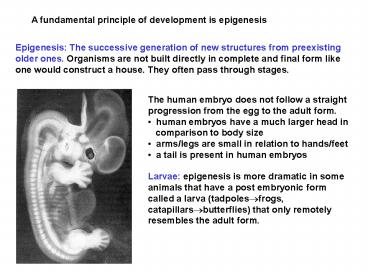
Epigenesis The successive generation of new
structures from preexisting older ones. Organisms
are not built directly in complete and final form
like one would construct a house. They often pass
through stages.
- The human embryo does not follow a straight
progression from the egg to the adult form. - human embryos have a much larger head in
- comparison to body size
- arms/legs are small in relation to hands/feet
- a tail is present in human embryos
Larvae epigenesis is more dramatic in some
animals that have a post embryonic form called a
larva (tadpoles?frogs, catapillars?butterflies)
that only remotely resembles the adult form.
2
Meiosis is the first step in gametogenesis
separation of homologous chromosomes into haploid
daughter cells
Spermatogonia and oogonia are the germ cells that
will eventually develop into the mature sperm or
egg Primary spermatocyte or oocyte the first
step in this development is the duplication of
homologous chromosomes to get ready for meiosis
Secondary spermatocyte or oocyte the first
meiotic division separates the homologous
chromosomes from each parent Spermatids or eggs
the second meiotic division separates the 2
chromatids and creates 4 haploid cells In males,
this eventually produces 4 sperm cells by the
process of spermiogenesis. In females, it
produces 1 egg and 3 polar bodies. This allows
the egg to retain more cytoplasm to support early
stages of development
3
The timing of meiosis differs in females and males
In males, the spermatogonia enter meiosis and
produce sperm from puberty until death. The
process of sperm production takes only a few
weeks. Each ejaculation has 100 to 500 million
sperm. In females, this process is more complex.
The first meiotic division starts before birth
but fails to proceed. It is eventually completed
about one month before ovulation in humans. In
humans, the second meiotic division occurs just
before the actual process of fertilization
occurs.
Thus, in females, the completion of meiosis can
be delayed for over 50 years. This is not always
good. Only I egg produced In addition, all
meiosis is ended in females at menopause.
4
Homologous chromosomes form the synaptonemal
complex which facilitates crossing over and
genetic diversity
During meiosis, homologous chromosomes join
together in pairs to form the synaptonemal
complex. Each pair of chromatids is connected by
axial proteins. The 2 homologous chromosomes are
held together closely by central element
proteins. A recombination nodule forms that
contains enzymes for cutting and splicing DNA.
Chromosomes are cut and joined crosswise at
points called chiasmata, seen when they
separate. The exchange of genetic material is
evident when the chromosomes separate This
process is dangerous as it leads to deletions and
duplications of genetic material. However, it is
also valuable because it increases genetic
diversity and facilitates evolution.
5
Spermatogenesis occurs in the seminiferous tubules
The mammalian testes are divided into many
lobules, and each lobule contains many tiny
seminiferous tubules. Sperm develop in an ordered
fashion in these tubules. Cells start to mature
on the outside and move inward (towards the
lumen) as the become mature sperm.
Spermatogonia are the most primative cells. They
differentiate as primary spermatocyte ? secondary
? spermatid ? sperm are released into
lumen. Sertoli cells are supporting cells that
stretch from the lumen to the edge of the tubule.
They nourish the developing sperm. They form a
blood-testis barrier to control spermatogenesis
(similar to the blood-brain barrier). These cells
also inhibit spermatogenesis before puberty and
stimulate the process after puberty.
6
Spermiogenesis is the maturation process into
sperm
The golgi vesicles combine to form an acrosomal
vesicle that lies over the nucleus. Its full of
enzymes
Centosomes start to organize microtubules into
long flagella
Mitochondria start to localize next to the
flagella to provide ready energy
The nucleus condenses in size and is stabilized
by special proteins called protamines
The excess cytoplasm is pinched off as a residual
body (no need for organelles and cytoplasmic
proteins)
Sperm are tiny, but highly specialized missiles
for delivering the male genome Microfilaments
shoot the acrosome into the egg to harpoon it
and pull it in. The acrosome has enzymes for
breaking into the egg. The midpiece has large
numbers of mitochondria for horsepower. The tail
has a powerful flagellum for driving the sperm
into the proximity of the egg (in humans, through
the uterus and up into the oviduct.
7
Spermatogonia and oogonia are stem cells
What is a stem cell? Stem cells have 3
properties 1. They are undifferentiated
cells 2. They have potential for
self renewal 3. They are able to
undergo differentiation to form committed
progenitor cells (a fancy
word for all types of differentiated
adult cells such as muscle, bone,
skin, etc)
8
Vitellogenesis is production of the major yolk
proteins
Yolk animal eggs contain large amounts of
protein, lipid, and glycogen to nourish the
embryo. These materials are collectively called
yolk. Yolk is minimal in animal eggs that
sustain only the first portion of embryogenesis
(humans and many mammals that have a placenta
need only support cleavage for several days
before implantation into the uterus). However,
yolk is stored in large amounts in the eggs of
birds and reptiles because their eggs have to
support the entire process of development.
Animal vegetal polarity In eggs that have a
lot of yolk, the yolk is concentrated in the
vegetal pole. The animal pole contains the
nucleus and relatively little yolk. The yolk in
the vegetal pole interferes with cytokinesis
leading to incomplete cleavage.
9
Fertilization in the sea urchin is divided into 5
steps
10
Microfilaments in sperm form the acrosomal
process which harpoons the egg and brings the
two together
When a sperm approaches the egg, an acrosomal
process forms and is activated. This is a
projection from the tip of the sperm. Microfilame
nts rapidly polymerize and extrude this at a
speed of 10 mm/second. Very rapid
polymerization. Light micrographs of
fertilization in the sea urchin taken at
intervals shown in seconds on the left. The sperm
is on the left, and the egg on the right. The
acrosomal process pentrates the egg envelopes and
pulls the egg into contact with the sperm.
Sea urchin
11
Bindin is the species-specific adhesive
material that binds sea urchin sperm to eggs
Bindin is a protein that is stored in the
acrosome. Bindin binds to receptors on eggs only
of the same species as the sperm.
Transmission electron microscopy showing bindin
to be localized on the sides of the acrosomal
process. It is also seen on the vitelline
envelope of the sea urchin egg. Sections
incubated with primary antibody to bindin and
subsequently with a secondary antibody linked to
electron dense material.
12
Plasma membrane contact triggers egg activation
Egg activation a series of events that cause a
quiescent egg to start mitosis. The process is
triggered by membrane contact and gamete
fusion A key mediator of egg activation is
phospholipase C, an enzyme found in the plasma
membrane of eggs. Phosholipase C activation
causes induction of 2 second messengers, DAG and
inositol triphosphate (IP3). Both stimulate the
activation of protein kinase C (IP3 acts
indirectly through Ca).
If phospholipase C is experimentally activated in
the absence of sperm, the egg still undergoes
activation. Although phospholipase C is
important, it does not appear to be necessary.
13
The fast block to polyspermy is caused by a
fertilization potential
The fast block starts as soon as 1 sec after egg
activation. The egg plasma membrane has a
resting potential of 75 mV due to differential
permeabilty to small ions (Na,
K). Fertilization potential immediately after
the sperm and egg touch, the membrane potential
changes to 20 mV, and remains here for 1 min.
The fertilization potential is induced by
depolarization of the membrane due to the flux of
different ions in different animals.
Sperm egg fusion and entry occur only in the
fleeting interval (1 sec) between 75 and 20 mV.
No more fusion with other sperm can occur after
the first second. This block lasts about 1 minute.
14
The cortical reaction assures a slow, long
lasting block to polyspermy
- The cortical reaction begins soon after egg-sperm
contact and is complete within 1 min. Ca and
protein kinase C interact to stimulate release of
cortical granules. The slow block is critical
because it is long lasting. - Cortical granules membrane bound vesicles
derived from the golgi and located directly
beneath the oocyte plasma membrane. They contain - Proteases that digest protein connections between
egg plasma membrane and vitelline envelope
(allows the two to separate). - Glycosaminoglycans that are released into the
perivitelline space and attract water, causing
this space to expand. - Peroxidase and other enzymes are released that
cross-link the vitelline envelope and make it
hard. - Enzymes remove receptor proteins that bind sperm.
15
What is cleavage?
- Cleavage is a rapid series of mitotic divisions
that occur just after fertilization. - There are two critical reasons why cleavage is so
important - Generation of a large number of cells that can
undergo differentiation and gastrulation to form
organs. - 2. Increase in the nucleus / cytoplasmic ratio.
Eggs need a lot of cytoplasm to support
embryogenesis. It is difficult or impossible for
one nucleus to support a huge cytoplasm, and
oocytes are one of the largest cells that exist.
One small nucleus just cannot transcribe enough
RNA to meet the needs of the huge cytoplasm. - A larger nucleus to cytoplasmic ratio is optimal
for cell function. Cell - division occurs rapidly after fertilization to
correct this problem.
16
Cleavage differs from normal mitoses in 2 respects
- Blastomeres do not grow in size between
successive cell divisions as they do in most
cells. This leads to a rapid increase in the
nucleus / cytoplasmic ratio. Cells undergoing
cleavage have mainly S and M phases of the cell
cycle (little or no G1 or G2). - Cleavage occurs very rapidly, and mitosis and
cytokinesis in each round of cell division are
complete within an hour. Typical somatic cells
divide much more slowly (several hours to days)
and even the fastest cancer cells divide much
slower than occurs in a zygote during cleavage. - Cleavage differs in different types of eggs. The
presence of large amounts of - yolk alters the cleavage pattern, leading to
incomplete cleavage that - characterizes birds and reptiles.
- Two areas of interest
- How does the process of cleavage differ in
different organisms? - What mechanisms regulate cleavage?
17
Eggs are classified by how much yolk is present
- Isolecithal eggs (iso equal) have a small
amount of yolk that is equally distributed in the
cytoplasm (most mammals have isolecithal eggs). - Mesolecithal eggs (meso middle) have a moderate
amount of yolk, and the yolk is present mainly in
the vegetal hemisphere (amphibians have
mesolecithal eggs). - Telolecithal eggs (telo end) have a large
amount of yolk that fills the cytoplasm, except
for a small area near the animal pole (fish,
reptiles, and birds). - Centrolecithal eggs have a lot of yolk that is
concentrated within the center of the cell
(insects and arthropods).
18
The pattern of cleavage of the zygote depends
upon the pattern of yolk distribution
- Holoblastic cleavage occurs in isolecithal eggs
(mammals, sea urchins). The entire egg is cleaved
during each division. - Meroblastic cleavage occurs when eggs have a lot
of yolk. The egg does not divide completely at
each division. Two types - a. Discoidal cleavage is limited to a small
disc of cytoplasm at the animal - pole. All of the yolk filled
cytoplasm fails to cleave (characteristic of - telolecithal eggs such as birds).
- b. Superficial cleavage is limited to a
thin surface area of cytoplasm that - covers the entire egg. The inside of
the egg that is filled with yolk fails - to cleave (centrolecithal eggs such
as insects).
19
Embryonic stem cells can be cultured from the
inner cell mass
Cells in the inner cell mass are
undifferentiated, they multiply indefinitely, and
are known as embryonic stem cells. Stem cells are
totipotent they have the potential to form any
tissue. These cells are of great scientific and
medical importance. They can be removed from the
embryo, genes can be introduced into the cells,
and then they can be placed back in the
blastocyst. This is how one constructs transgenic
or knock out mice. The embryonic stem cells are
also used to grow certain types of tissue in
culture. Theoretically, it should be possible to
grow structures such as ears, muscles, nerves,
and skin for transplantation to sick
individuals. Interestingly, if you inject adult,
differentiated cells back into the environment of
the morula or blastula, they become
undifferentiated, and they can redifferentiate to
form many parts of the body.
20
The mitotic spindle determines the orientation of
the cleavage plane
Blastomeres can cleave either equatorially or
meridionally. Cytokinesis usually directly
follows mitosis, except for superficial cleavage.
Cytokinesis invariably occurs in a plane
perpendicular to the axis of the mitotic spindle.
Thus, the spindle orientation controls the
orientation of the contractile ring The proximity
between the egg cortex and the mitotic spindle is
also important for furrow formation. In eggs
where the the outer cortex is displaced from the
spindle (birds and insects), by large amounts of
yolk, the spindle never activates the cleavage
furrow.
How does a blastomere know to divide meridionally
or equatorially?
21
Mitotic spindles are oriented with their
axis parallel to the longest available cell
dimension
Mitotic spindles work to keep the cell round in
shape. Experiment It is possible to control how
tightly blastomeres adhere by changing the
concentration of calcium. High calcium
concentrations cause more cell cell attachment.
Low calcium causes minimal attachment. The effect
is likely mediated by adhesion molecules such as
cadherin.
When blastomeres adhere they have a longer axis,
and the mitotic spindle is almost always oriented
parallel to this axis. As the cell becomes more
spherical in low calcium medium, the mitotic
spindle orientation starts to become random.
22
How does a cell know when it should divide?
The cyclic activity of a protein dimer controls
the activity of the cell cycle Cyclin dependent
kinase 1 (cdk1) is an enzyme that is always
present in cells. It can phosphorylate other
proteins when it is activated. Cyclins are a
family of proteins that are produced in cyclic
fashion during the cell cycle. Cyclin B is
destroyed shortly after metaphase, but
accumulates slowly thereafter.
M phase promoting factor (MPF) when there is
sufficient cyclin B, it combines with cdk1.
Additional regulatory changes occur such as
phosphorylation of threonine and
dephosphorylation of tyrosine. The active
kinase phosphorylates specific cell proteins that
control mitosis (spindle, nuclear lamins, and
chromosomes). The actual targets of M phase
promoting factor are an area of intense research
interest.
23
Timing of cleavage divisions
Normal eukaryotic cells divide slowly, once every
several hours or days. The cell cycle has G1 and
G2 periods. During G1 the cell synthesizes RNA
and other components for cell growth. Cleavage
consists of very rapid successive mitoses. Since
the egg has stored large amounts of RNA and other
material, it does not need G1 or G2. However, as
the number of cells increases, the nucleus /
cytoplasmic ratio also increases. The rate of
cell division slows because the cell now needs to
synthesize its own RNA and grow between
divisions. Thus, G1 and G2 are restored
midblastula transition.
24
How is cell differentiation regulated how does a
cell know what to do?
There are at least 3 theories to explain cell
differentiation Chromatin diminution
originally hypothesized in the 1800s. Cells enter
a specific differentiation pathway because
selected genes that control other pathways are
lost. Out of favor, although a few concrete
examples exist. Selective gene amplification
Differentiation is due to selective amplification
of a small set of genes that induce one type of
differentiation. For example, liver cells would
have amplification of liver specific genes.
Genomic equivalence all cells in an organism
have complete and equivalent sets of genes.
Differentiation is caused by differential
expression of these genes. Evidence for this
theory is strong in plants, but less in animals.
25
Nuclei from embryonic cells are still totipotent
Nuclear transfer the most direct way to test for
totipotency of the nucleus is to transplant it
into an egg of the same species. This has been
done with frog eggs (Rana pipiens). Using a fine
glass needle, the egg nucleus is sucked out.
Donor cells were taken from the frog embryo at a
later stage of development. The cytoplasm was
smashed, but the nucleus was saved and injected
into the egg. This technique was used to produce
tadpoles and some normal frogs. Similar
experiments were performed successfully with
Drosophila eggs. However, nuclei were competent
only until the gastrula stage.
26
Nuclei from older donors show decreasing ability
to promote development
When nuclear transfer experiments were extended
to older cells, there was an age dependent
decrease in success. In Rana pipiens, nuclei
isolated from tail bud tadpoles could no longer
induce development. In Xenopus laevis, it worked
a little better, but most tadpole nuclei still
did not work.
Why doesnt it work in animals like it does in
plants? Older nuclei start to lose genetic
information? Some other technical
problem? Spermatogonia from adult frogs are
clearly totipotent, but they also did not work
well in nuclear transfer experiments.
27
Nuclei from mature differentiated cells are not
prepared for the rapid mitotic cycles of cleavage
Mature adult cells divide at a slow pace (days to
weeks between divisions). When adult nuclei are
placed into eggs, they just can not keep up with
the rapid pace of DNA replication.
These nuclei quickly develop DNA breaks that are
caused by abnormal mitoses. Only about 60 of
the adult nuclei have replicated their DNA before
the first mitotic division. Nuclei isolated from
zygotes replicate DNA much more rapidly.
28
Sheep can be cloned by fusing adult cells with
enucleated eggs nuclei from mature differentiated
cells are not prepared for the rapid mitotic
cycles of cleavage
- Recently, better results have been obtained using
sheep. Several technical changes may have
contributed to this success. - Used only donor cells from the G1
- phase of the cell cycle
- fused the whole cell with an
- enucleated egg
- The slowly dividing nucleus appears to be better
able to be reprogrammed for rapid cell division. - Used 2 distinct strains of sheep so that the
identity of offspring would be clear. - In one of 277 fused oocytes, a completely normal
sheep developed from a 6 year old breast
epithelial cell.
29
Embryonic cells are broadly classed as epithelial
or mesechymal
Epithelial cells are well-differentiated. They
compose skin and line the body cavities (ie, the
digestive tract). They are polarized. Their
apical surface faces out and their basal surface
rests on the basement membrane (extracellular
matrix that supports cells). Epithelial cells are
closely connected with adjacent cells by
specialized attachments including tight
junctions, gap junctions, and desmosomes.
Mesenchymal cells are poorly differentiated and
have the potential to develop into many different
tissues, including epithelial cells. They have a
leading edge with lamellipodia, and a trailing
edge. They are not connected to adjacent cells
but they are in contact with the extracellular
matrix.
30
Vegetal plate the first step in sea urchin
gastrulation is formation of the vegetal plate, a
thickening of epithelial cells in the vegetal
pole.
Primary mesenchymal cells these cells change
adhesive properties and the large micromeres
start to migrate into the blastocoel as free
mesenchymal cells (ingression). Mesenchymal cells
are loose cells that can differentiate into many
different organs. Archenteron the primitive
gut. The archenteron is formed in several stages.
1. the vegetal plate invaginates into the
blastocoel, 2. It elongates by convergent
extension. 3. It hooks up with the front and is
pulled forward, and 4. Involution occurs with
movement of cells around the blastopore and into
the archenteron. What forces drive the process
of gastrulation?
31
The primitive groove and pit are the site of
gastrulation in birds
Crossection of blastoderm (blastula in
birds). Epiblast is the upper layer of epithelial
cells, blastocoel is the space below the
epiblast, and hypoblast is the lower layer of
epithelial cells.
Epiblast cells roll over the primitive ridge and
involute into the groove. The cells lose contact
with one another and migrate inwards by
ingression Mesoderm. 3 germ layers are
established
32
Gastrulation in a 16 day old human embryo
- The primitive streak and Hensons node form just
as in birds. - The cell movements are similar
- Cells roll over the primitive ridge and into the
groove - They ingress individually and move out to form
discs with the 3 germ layers - The 3 layers also move laterally to form the
extra embryonic endoderm and - mesoderm even though there is no yolk to
digest. This is surprising - because mammalian embryos could gastrulate
easily by invagination as sea - urchins (they have a placenta and no yolk).
- Instead, gastrulation appears to recapitulate a
pattern established by bird- - like ancestors and reptiles.
33
Molecular control of gastrulation and
morphogenesis
Does each cell in the blastula have detailed
instructions in the DNA that tell it exactly
where to go during gastrulation?
If an embryo is disaggregated into individual
cells, each should know exactly where to go to
reform a new embryo. When this experiment is
performed, a degree of reorganization occurs, but
it is not complete. Embryoids are slightly
similar to embryos but they lack the real
organization. Conclusions 1. Genes impart only
partial instructions for assembly of the
embryo 2. Like cells all stick together,
revealing distinct adhesive properties. 3. The
relative positions of aggregates reflect the
relative positions in the embryo (skin outside,
heart inside).
34
Cell adhesion is the driving force in gastrulation
When cells from an embryo are disaggregated and
recombined, they can be readily ranked according
to their ability to form the central
portion. (chondrocytes gt heart cells gt liver
cells is the hierarchical order)
Why? Differential adhesion hypothesis the cell
type with maximal adhesiveness (chondrocytes)
will form a core that is surrounded by concentric
spheres of cells with progressively lower
adhesiveness. Cell adhesion can be measured by
the pancake test. When aggregates of different
cell types are subjected to a flattening force
(centrifugation to induce a centrifugal force),
the cells that adhere most tightly form a ball,
while those that adhere more loosely form a
flatter, pancake structure. Cell adhesion is a
major factor that regulates aggregation of like
cells and controls position during morphogenesis.
What regulates how tightly or loosely cells
attach?
35
Cells adhere by cell junctions, cell adhesion
molecules, or substrate adhesion molecules
Cell junctions large, complex structures that
form slowly but generate very strong and durable
connections (tight junctions, desmosomes, and gap
junctions). Cell adhesion molecules (CAMs)
single molecules that traverse cell membranes and
allow cells to adhere to one another. Adhesions
form quickly, they are selective, but they are
relatively weak in comparison to cell
junctions. Substrate adhesion molecules (SAMs)
a group that consists of extracellular matrix
molecules and matched receptors that are
expressed on the cell surface.
36
CAMs firmly anchor adjacent cells to the
cytoskeleton
Cell adhesion molecules (CAMs) are glycoproteins
with 3 major domains The extracellular domain
allows one CAM to bind to another on an adjacent
cell. The binding can be to the same type of cell
(homotypic) or to a different cell type
(heterotypic). The transmembrane domain links
the CAM to the plasma membrane through
hydrophobic forces. The cytoplasmic domain is
directly connected to the cytoskeleton by linker
proteins. This anchoring is important to prevent
lateral diffusion of adhesion molecules in the
membrane.
Three major types of CAMs are immuno
globulin-like CAMs, cadherins, and lectins.
37
CAM expression during gastrulation is correlated
with cell fate
Fate map it is possible to predict which parts
of the blastula will develop into specific
structures after gastrulation. Expression map of
CAMs it is possible to localize expression of
CAMs using in situ hybridization and
immunostaining of the blastula. Cells with
different fates express different CAMs. Cells
destined to become neural tissue express high
levels of N-CAM. Cells destined for epidermis
express E-cadherin.
The respective cell adhesion molecules are
expressed before the cells actively start to form
the adult tissue. This suggests that CAM
expression is important in fate
determination. During gastrulation, cells go
where their CAMs lead them
38
Changes in cell adhesion are important for
gastrulation
Gastrulation in the sea urchin is initiated by
specific changes in cell adhesion. One of the
first steps is ingression of mesenchymal cells
from the vegetal plate into the blastocoel to
form the skeleton of spicules. The mesenchymal
cells lose their adhesion to hyaline and the
adjacent nonmesenchymal blastomeres. They start
to increase adhesion to the basement membrane and
material within the blastocoel. These changes can
be measured by isolating specific cells and
testing adhesion in culture. E-cadherin is lost
from the ingressing cells due to endocytosis of
specific areas where it was expressed. Levels of
b-catenin are also reduced on these cells.
39
Fibrous ECM components provide contact
guidance to migrating cells during gastrulation
The movement of cells during gastrulation may
also depend upon expression of ECM. ECM allows
migrating cells to attach transiently while
moving over the surface. During gastrulation in
amphibians, cells move into the blastocoel and
migrate over the inside of the roof. If a
portion of the roof is cut out and inverted, no
movement of gastrulating cells occurs here. This
suggests that some CAMs or SAMs may be
missing. What molecules would this be?
40
Fibronectin on the inner roof of the
blastocoel is critical for gastrulation
Immunostaining of the blastocoel shows that
fibronectin was expressed in abundance on the
inner roof. Fibronectin binds to integrins on the
membrane. Neutralizing antibody to fibronectin
was injected into the blastocoel to test the role
of fibronectin. This aborted gastrulation. Since
no epidermal cells could migrate into the
blastopore, many cells accumulated on the
surface, forming deep folds. If an unrelated
antibody was injected, there was no
inhibition. Fibronectin binds integrins through
an RGD sequence. Similar results were obtained by
injecting the tripeptide RGD. Furthermore,
blocking the integrin receptor with injected
antibodies also inhibited gastrulation.
Fibronectin is important for contact guidance of
migrating cells during gastrulation.
41
PC12 cells resemble chromaffin cells which can
differentiate into neurons. When they convert to
the neural phenotype they express N-CAM and
N-cadherin on their cell surface. When PC12
cells are grown on cells that do not express
N-CAM or N-cadherin (3T3 cells) they retain the
undifferentiated chromaffin phenotype.
If the PC12 cells are grown on the same cells
that have been transfected with N-CAM or
N-cadherin genes, they convert to the neuronal
phenotype. They form long dendrites and express
neuronal genes. Differentiation is accompanied
by opening of calcium channels
42
Histogenesis is the process by which cells and
tissues acquire functional specialization
In humans Cleavage ? gastrulation ?
organogenesis ? histogenesis (2 weeks) (1
week) (4 weeks) (7
months) Organogenesis the formation of organ
rudiments to establish the basic body
plan. Histogenesis differentiation of cells
within the organs to form specialized tissues.
Tissues are composed of cells and extracellular
material that perform a specific function. Each
specific tissue develops mainly from one germ
layer.
embryo
fetus
43
Spemanns famous organizer experiment
Spemann originally thought that the donor dorsal
lip of the blastpore differentiated into neural
tube and structures of the embryonic axis. To
prove this, he grafted dorsal lip blastopore
tissue from a nonpigmented donor newt onto
another gastrula from a heavily pigmented
species. When the embryos developed he found that
the embryonic axis was actually composed of
pigmented recipient tissue. Only a small strip
of donor tissue was present in the middle of the
neural plate.
44
Spemanns experiments were confirmed in birds
- 3 conclusions about the organizer
- The dorsal lip of the blastopore developed
according to its own fate. It gastrulated
normally in the new location and formed
notochord. - The graft dorsalized the hosts ventral mesoderm
(converted gut tissue to kidneys and somites). - The graft acted as a neural inducer. It caused
host ectoderm to form a neural plate and close to
become a neural tube.
45
Axis induction by disinhibition
Early experiments by Spemann studied how the
organizer worked. If the cells of the dorsal lip
were killed or crushed, the activity was still
present. Activity could be mimicked partially by
changes in pH or ionic strength. With the advent
of molecular biology 2 areas have been
explored 1. Induction certain gene products
directly induce neurulation in either dorsal or
ventral ectoderm. Ventral ectoderm doesnt get
enough.
2. Disinhibition the normal pattern for
ectodermal development is neurulation. Ventral
ectoderm escapes from this path by producing an
inhibitor of neurulation. The organizer works by
inhibiting the inhibitor. Cells cultured from
ventral ectoderm differentiate as neurons only if
cultures are sparse. Bone morhogenetic protein
(BMP-4) is released by ventral ectoderm cells. It
stimulates formation of ventral structures and
inhibits neurulation if injected into
embryos. If BMP-4 is blocked, dorsal structures
replace ventral structures (dominant negative
mutants or innactivate the BMP-4 receptor).
46
Spemanns organizer (dorsal lip) inactivates BMP-4
Chordin and noggin the dorsal lip of the
blastopore produces 2 proteins that antagonize
the action of BMP-4. Chordin and noggin each bind
to BMP-4 and prevent it from binding to its
receptor. Chordin and noggin have strong
dorsalizing effects. They cause excessive head
development and block ventral differentiation if
injected into the embryo.
What turns on chordin and noggin? Goosecoid may
be the master regulator that is similar to
Spemanns organizer. Expression of goosecoid is
limited to the dorsal lip of the blastopore. It
is turned on at the right place at the right
time. Goosecoid encodes a transcription factor
that can activate chordin gene expression. Work
is continuing to explore how this gene functions.
47
(No Transcript)
48
The notochord and floor plate induce
the dorsoventral pattern of neural development
To examine whether ventral columns were induced
by adjacent tissue, an extra notochord was
grafted to the side of the developing neural
plate.
This created an extra floor plate and ventral
column, suggesting that it induced these
structures. If the notochord was removed, no
ventral column or floor plate developed,
consistent with the above idea. It is believed
that the notochord first induces the floor plate
and the floor plate then induces ventral columns.
This was confirmed by grafting a floor plate,
which also induced efferent nerves. What is the
molecular nature of this inducer of dorsoventral
patterning?
49
Sonic hedgehog (shh) induces the dorsoventral
pattern
Sonic hedgehog (shh) is a gene that is expressed
in the notochord at first and later in the floor
plate. Mice that lack shh fail to develop floor
plates in the CNS. Shh is a secreted
glycoprotein that induces a gradient that is high
near the floor plate and progressively lower in
dorsal regions. Shh initially induces neural
plate cells to form floor plate. Other signals
from the dorsal ectoderm direct the dorsal
columns and the roof plate. Different levels of
shh appear to specify different types of neuron
differentiation.
50
Neural crest cells form a variety of tissues
The fate of neural crest cells has been mapped by
a number of techniques (radioactive tracers,
transplants from pigmented species to albinos).
There are 2 patterns of migration in the trunk
region Dorsolateral path enter skin and form
melanocytes Ventral path form afferent neurons
of dorsal route ganglia, sympathetic and
parasympathetic ganglia, and adrenal medulla
Neural crest cells help to form addition
structures in the head such as bones, connective
tissue, eyes, ears, and teeth. They also help
to form blood vessels and connective tissue in
the trunk
51
How do neural crest cells differentiate into many
tissues?
Pluripotency hypothesis each neural crest cell
has the potential to form any or all structures.
Inductive signals from adjacent tissue determines
their fate. Selection hypothesis the neural
crest contains a mixed population of
predetermined cells. Each cell has only one
possible fate and it migrates according to this
fate.
The real truth may lie between these two
extremes. Clonal analysis when individual
neural crest cells are placed in culture, it is
clear that a single cell can give rise to others
that differentiate into multiple cell types
(pigment cells and neurons). Premigratory cells
have a wider potential than do the cells that
have already started to migrate. They may become
partially differentiated as they migrate.
52
Formation of the eye involves reciprocal
interactions
Lens placode the ectoderm invaginates in
response to signals from the optic cup
underneath. It then pinches off as a lens
vesicle. Cells elongate to fill the vesicle and
start to synthesize crystallins. Optic cup
forms from the neural tube by invagination. The
opening (choroid fissure) closes forming a round
optic cup, an extension of the brain. Optic
stalk connection to the brain that is filled
with neurons to form the optic nerve. Reciprocal
interaction the lens induces the formation of
the optic cup and the cup regulates formation of
the lens. When the lens from a species with large
eyes is transplanted, it induces an extra large
optic cup and it also does not grow as large as
usual.
53
Mammary gland development in humans mimicks
ancestral patterns
In normal development of humans, only one pair of
segments in the ridge survives, and the remaining
precursors degenerate. In some individuals, the
other segments of the mammary ridge fail to
degenerate, so that accessory nipples or breasts
are formed.
Primates evolved from small creatures that nursed
multiple offspring. An extended mammary ridge
would have given human ancestors a survival
advantage. Two breasts are obviously more
adaptive for humans who normally have only one
offspring at a time. Atavism the occasional and
abnormal persistance of a primitive adult feature
in an evolved species (multiple mammary glands).
It is easier to modify an older pattern of
development than to develop a totally new
pattern. This idea is a pervasive in
developmental biology.
54
Histogenesis of organs derived from mesoderm and
endoderm
Endoderm endodermal derivatives consist of the
lining of the digestive tract and derivatives.
These include liver, pancreas, bladder, lungs,
and thyroid. Mesoderm a wide variety of tissues
including bone, muscle, urogenital organs
(oviducts, uterus, epididymis), circulatory
system (blood and vessels), connective tissue,
kidney, and heart. Histogenesis of endodermal
and mesodermal tissues provide examples of two
recurrent themes in developmental
biology Recapitulation the occurrence of a
phylotypic stage in development at which all
species of a phylogenetic group show an uncanny
similarity. Advanced organisms, such as humans,
often go through developmental stages that
resemble more primitive ancestors (birds or
fish). Reciprocal interaction many organs
originate from two embryonic rudiments, usually
one is epithelial and one mesenchymal. During
development, the rudiments exchange signals to
insure that both structures develop harmoniously
(lung and kidney)
55
Craniocaudal flexion also aids in closing the gut
Craniocaudal flexion rapid extension of the
neural plate just before closing causes the
anterior and posterior regions to flex
downward. Vitelline duct the wide opening to
the yolk sac becomes a narrow passage. Buccophary
ngeal membrane forms at the anterior end of the
gut. Cloacal membrane forms at the caudal
end. Pharynx the gut between the bucco
pharyngeal membrane and the lung bud. Foregut
the gut from the lung bud to the rudiment of the
liver and pancreas. Midgut/hindgut from the
liver to the cloacal membrane. Bending inward at
the lateral boundaries and from the anterior and
posterior is involved in initiating formation of
gut structures
56
The embryonic pharynx contains a series of arches
The development of the pharynx in humans and
other mammals actually recapitulates that of more
primitive ancestors (birds and fishes). Pharyngea
l pouch bulges in the pharyngeal endoderm that
push away the mesoderm. This induces the
formation of an overlying pharyngeal cleft in the
ectoderm. In some species, the pouches open to
form gill slits between the arches. The
pharyngeal arches were probably used by primitive
ancestors to filter food from the water. In fish,
they have developed into gills.
57
The phylotypic stage of development occurs in all
species of a phylum
Recapitulation advanced vertbrates recapitulate
embryonic stages of their phylogenetic ancestors.
After the phylotypic stage, each species displays
a specialized pattern of differentiation. Why??
you once looked like a fish!
Evolution seems to modify existing patterns of
development rather than creating new ones. It is
speculated that mutation of genes that control
the phylotypic stage may prove lethal. Changing
is easier than reinventing.
58
The mesoderm organizes into four major regions
The mesoderm lies between the sheets of
ectodermal and endodermal epithelia. It can form
epithelial structures, however, it frequently
forms mesenchyme, isolated cells surrounded by
extracellular matrix. The mesoderm organizes
into four major regions called axial, paraxial,
intermediate, and lateral plate mesoderm.
Axial mesoderm located along the dorsal midline.
It forms prechordal plate and cranial cartilage
in the head. It forms notochord in neck, trunk,
and tail. The notochord is a long rod of
connective tissue that is replaced by the
vertebral column during embryogenesis. Notochord
forms intervertebral discs.
59
Each section of the somite forms specific
structures
The fate of cells within the somite has been
traced by labeling cells with dye or
radioactivity. It forms 4 structures
1. Sclerotome cells from the ventromedial
portion become mesenchymal and divide rapidly.
They migrate to surround the notochord and neural
tube. They form cartilage which later becomes the
vertebral column and ribs. Dermomyotome this is
what remains of the somite after the sclerotome
leaves. 2. Dermotome the outer layer of cells
migrates dorsally to form the connective tissue
below the epidermis dermis. 3. Epaxial
myotome a lip formed at the dorsomedial margin.
These cells form muscles of the dorsal trunk. 4.
Hypaxial myotome the lip at the ventrolateral
margin migrates ventrally to form the muscles of
the limbs and ventral trunk.
60
Model of transverse patterning in somites
Ventralizing signals caused by sonic hedgehog
induce the sclerotome, which forms the vertebral
column and ribs. Dorsalizing signals induced by
the Wnt family/goosecoid contribute to induction
of epaxial myotome and dorsal trunk
muscles. Lateralizing signals due to BMP-4 may
cause the hypaxial myotome to form, producing
ventral trunk and limb muscles.
61
Intermediate mesoderm forms the kidney
The intermediate mesoderm is located between the
somites and lateral plates.
Kidney the principle function is to eliminate
waste products. Blood is filtered through the
kidney, nutrients and serum proteins are
reabsorbed from the filtrate, and waste products
(urea) are excreted. Recapitulation Three types
of kidneys develop in vertebrates Pronephros
(probefore, nephroskidney) formed in the neck
region of all vertebrate embryos, but persists to
adulthood only in some fishes. Mesonephros
(mesomiddle) formed for much of the length of
the trunk in most vertebrate embryos. It persists
in most fishes and amphibians. Metanephros
(metaafter) the metanephros forms in the
posterior and becomes the kidneys in birds,
reptiles, and mammals.
62
Reciprocal interactions are important for
development of the metanephros
Step 1 the metanephrogenic mesenchyme induces
the adjacent mesonephric duct to form the uteric
bud. The bud is stimulated to enter the
mesenchyme and branch to form collecting ducts.
Polypeptide growth factors are secreted by the
mesenchyme and activate the growth of the uteric
bud. Step 2 the developing bud stimulates the
mesenchyme to form the nephrons (gt 1 million /
kidney in humans). The mechanism of induction is
unclear. E-cadherin increases, allowing cells to
adhere. Increased expression of laminin and
collagen IV.
63
Development of the heart
Recapitulation in heart development heart
formation in humans and higher vertebrates
recapitulates early stages of development in
fish. The heart develops from bilateral
rudiments in the visceral layer of mesoderm.
These rudiments fuse to form a primitive heart
tube. Endocardium the inner layer is continuous
with the endothelium of blood vessels. Cardiac
jelly a thick layer of extracellular matrix
surrounding the endocardium. It facilitates cell
movement as the endocardium develops. Myocardium
visceral mesoderm forms the heart muscle.
64
The embryonic heart tube is divided into 4
chambers sinus venosus, atrium, ventricle, and
truncus arteriosus. This primitive arrangement is
the final form in many fishes. Early human
embryos look almost identical.
The embryonic heart tube undergoes dramatic
changes in mammals. The interatrial and
interventricular septa arise. The
atrioventricular connection is remodeled so that
2 valves form. The truncus arteriosus splits to
form the aorta and the pulmonary artery.
Splitting is accomplished by forming ridges of
tissue that grow together in each tube.
65
Extraembryonic membranes
Fish and amphibian embryos develop in water which
provides several advantages a source of food,
protection against trauma, dessication, and a
reservoir for excreted waste. Reptiles, birds,
and mammals lay eggs on land and have developed
extraembryonic membranes to assist with these
developmental processes. There are 4 membranes
amnion and chorion develop from somatopleure, a
bilayer of ectoderm and somatic mesoderm. The
allantois and yolk sac are from splanchnopleure,
a bilayer of endoderm and visceral mesoderm.
66
How do the membranes function in reptiles and
birds?
Amnion a sac filled with fluid allows the embryo
to float , shock absorber Chorion the chorion
and allantoic membranes form blood vessels that
enable gas exchange with the embryo, embryonic
lung Allantois also forms a reservoir for
metabolic wastes, embryonic kidney Yolk sac
forms blood vessels to carry nutrients from the
yolk to the embryo proper, embryonic digestive
system
67
What happens to these membranes in mammals?
The same membranes form in mammals, however, the
functions are further modified. Mammals develop
in the uterus rather than an egg. Amnion
continues to surround and protect the
embryo. Chorion forms the cytotrophoblast that
makes chorionic villi and forms the fetal portion
of the placenta. Yolk sac this is formed even
though there is no yolk. It will become the site
for germ cell formation (sperm and eggs). An
example of recapitulation.
68
Saturation mutagenesis
Saturation mutagenesis method for developing
mutations in all genes that effect a specific
trait. Large numbers of organisms are mutagenized
and then bred. Offspring are carefully screened
for changes in a particular trait. Loss of
function mutants lessen the function of a gene
(most common) Null mutants completely lose
function Gain of function mutants activate gene
in the wrong place or the wrong time Features
that are necessary for saturation mutagenesis are
small numbers of genes, organisms that are cheap
and easy to breed and maintain, short gestation,
tolerant of mutagenesis. Drosophila and C.
elegans have been used frequently for this work.
Mice are not appropriate due to the longer
gestation time, expense, smaller litter size, and
larger genome size.
69
3. Homologous recombination to create a knockout
(null mutation) a cloned gene is prepared with
one or two exons replaced by an antibiotic
resistance gene (neo). The flanking homologous
sequences are retained. When introduced into
cultured stem cells, the transgene recombines
with the normal gene and knocks out its function.
This occurs in only a few cells but these can be
selected by growing the culture in antibiotic.
The purified stem cells are injected into the
inner cell mass of another mouse, and the embryo
is transplanted to a surrogate mother. Some of
the resulting chimeric mice should express the
transgene in the germ line. Since only one allele
is knocked out, you cross 2 heterozygotes and
hope for 25 homozygous offspring. In practice,
you often get none because the knocked out gene
was important for embryonic development.
70
Hox genes and pattern development of vertebrates
Pattern formation harmonious arrays of different
elements, such as the array of fingers on the
hand, body pattern (head, trunk, and tail), or
limb patterns. Pattern formation is best
understood in Drosophila, where most genes that
contribute to the body plan are described.
Anteroposterior axis in vertebrates is specified
by a group of genes called homeobox genes. There
are many similarities to Drosophila. The
dorsoventral axis in vertebrates is also
specified by genes that have counterparts in
Drosophila. Interestingly, although vertebrates
and invertebrates share a similar body plan, it
is inverted. Vertebrate limb development is
controlled by multiple genes, including homeobox
genes.
71
Cre lox recombination system
A new technology can overcome the problem of
blocked development. It allows one to study what
happens when the gene is suddenly knocked out in
adult cells (similar to a mutation that might
contribute to cancer). It is useful for studying
many important genes that are required for
embryonic development. Cre a recombinase
isolated from bacteriophage lambda. It cleaves at
all sites in a DNA sequence that have a specific
lox site. Construct a cre gene driven by an
inducible promoter such as metallothionine.
Lox genetically engineer the gene to be
knocked out such that it is flanked by lox
sites. Make one transgenic mouse for cre and
another for lox. Breed the mice so that one pair
is homozygous for lox and also has a cre gene.
When these mice are treated with zinc, the
metallothionine promoter is turned on and the cre
recombinase that is produced in all cells cuts
out the transgene flanked by lox sites. This
creates a knockout in the adult mouse that would
not be possible with conventional methods.
72
Hox genes specify anteroposterior body pattern
The physical order of Hox genes within the
complex is related to their order of expression
along the anteroposterior axis of the
embryo!! Genes at the 3 end are expressed in
the anterior and genes at the 5 end are
expressed progressively further
posteriorly. Hoxb cluster is expressed in the
central nervous system. Each gene is first
expressed at a sharply defined point and
expression continues posteriorly and gradually
tapers off. Rhombomeres the pattern of Hox gene
expression often coincide with repetitive bulges
in the sides of the rhomencephalon.
mouse
73
What accounts for the pattern of Hox gene
expression?
mouse
Enhancer sharing the sequential order of Hox
genes within the complex may result from the fact
that all genes in the complex share a common
enhancer element. If any gene is removed, or if
the complex is broken apart, the removed genes
may not be expressed properly. Thus, the
sequential order has been preserved during
evolution of different organisms.
74
Hox gene expression contributes to limb
development
Limb buds the first stage of limb development
occurs at 5 weeks in humans when small paddle
shaped limb buds form. Apical ectodermal ridge
(AER) a specialized structure formed by the
ectodermal covering of the limb bud. It is a
ridge that runs anterior to posterior. Progress
zone underneath the AER lies a zone of
mesenchyme that actively proliferates to form the
limb. Somites contribute mesenchyme to form
muscles and lateral plate mesoderm forms
cartilage and connective tissue.
The limb is formed by differential growth of
mesenchyme cells, by programmed cell death
(between digits), and specific patterns of
differentiation induced by Hox genes and other
factors. How does the limb know where to form?
How do arms become different than legs? How does
the limb develop its three axis?
75
Limb position is determined by FGF and Hox genes
The position of limb development depends on
signals from other tissues. Fibroblast growth
factor (FGF) these growth factors are produced
by mesenchyme (FGF-10) and epidermis (FGF-8) to
induce limb formation. If a small bead containing
FGF is implanted under the skin, an extra limb
develops. If the bead is implanted in the flank
near the anterior, it forms a wing. If the bead
is implanted posteriorly, it forms a leg. Knock
out mice lacking FGF-10 fail to develop limbs and
have no apical ectodermal ridge or zone of
polarizing activity.
76
Reciprocal interaction is critical for wing
development
Interactions occur between the AER on the surface
and limb bud mesenchyme that lies underneath in
the progress zone. Limb bud mesenchyme induces
formation of the AER from limb bud ectoderm. If
the ectoderm of a limb bud is removed at an early
stage of development, the mesoderm induces a new
AER. If the AER is removed during a later stage,
the limb mesenchyme stops growing and the limb is
truncated. If the limb bud mesenchyme is removed
or replaced with other tissue, the AER quickly
degenerates. The AER serves a permissive rather
than an instructive role. If you reverse its
orientation, digits will develop normally. If you
replace wing AER with leg AER, the wing develops
normally. The mesenchyme is the instructive
influence for limb development.
77
Caloric restriction postpones senescence
One of the most reliable ways to prolong life in
laboratory animals is to simply restrict their
calories. When rats were maintained on a low
calorie diet throughout life, they were 15
smaller but they lived 50 longer than litter
mates that ate ad libitum (all they wanted). If
restriction of calories is started later in life,
it still works, but lifespan is only extended
20. Food restricted rats show less evidence of
cancer, atherosclerosis, and autoimmune disease.
Why does restriction of calories delay
senescence? The underlying mechanisms are being
studied, but are currently unclear. Caloric
restriction induces levels of some antioxidant
enzymes.
78
Oxidative damage hastens senescence
Oxidative phosphorylation the process of
producing energy through conversion of O2 to H20
and creation of ATP. Although this reaction is
necessary for survival, it also produces
hazardous by products. Reactive oxygen species
(ROS) several highly reactive radicals are
produced at potentially damaging levels during
oxidative phosphorylation (superoxide radical,
hydroxyl radical, and hydrogen peroxide). Detoxif
ication enzymes several enzymes assist in
detoxification of ROS
Antioxidants natural molecules that reduce
oxidative damage from free radicals. These
include glutathione, vitamin C, vitamin E, and
b-carotine.
79
Telomerase and senescence
Somatic cells of humans show a limited capacity
for proliferation in cell culture, and this
corresponds to aging within the organism.
Hayflicks exp. Phase 1 cells grow rapidly when
placed in culture (30-60 pd) Phase 2 cell growth
rate slows after many population doublings Phase
3 cells stop growing and can never again enter
the cell cycle. However, cells
remain viable for extended periods.
The number of population doublings that occurs
before cells become senescent varies in cells
from different tissues. Cells from young
individuals divide many more times than cells
from old individuals. This suggests that all
somatic cells are capable of a limited number of
divisions. Thus, cell senescence may contribute
to organismal senescence.
80
Telomeres and senescence
One trigger for cell senescence is shortening of
telomeres. Telomeres are short repeats of DNA
(GGGTTA in humans) that form caps at the ends of
chromosomes to protect the ends from wearing
down. Each time that a cells divides, it loses
about 100 bp of telomeric DNA. Telomeres are
about 10 kbp long in embryonic cells. After about
80 cell divisions, telomeres wear down to 2 kbp,
which is thought to be a minimum length that
triggers senescence. Telomerase a reverse
transcriptase that restores telomeres. The enzyme
is active in germ cells and stem cells, but
activity is not detected in most somatic cells.
Telomerase is reactivated in cancer cells or
immortal cell lines.
81
Cell Death apoptosis versus necrosis
Cell death occurs in two ways Necrosis occurs
in response to injury. Cells are lysed and
release their contents. The membranes break down
causing release of organelles , DNA, and
lysosomes to interact with the adjacent cells.
This induces damage and inflammation (heart
attack, bruise). Apoptosis or programmed cell
death is when the cell commits suicide for the
good of the organism. Most cells have an
intrinsic cell death program that must be
constantly suppressed by survival factors
(cytokines and growth factors). Apoptosis is a
genetically controlled event, it requires energy,
and it allows cell death to occur in a very
controlled manner. The nuclei shrinks and
fragments into small pieces. These are easily
phagocytized by macrophages. There is no damage
to adjacent cells and no inflammation.
Recommended
CrystalGraphics Presentations





























