
From: Mathews Van Holde - PowerPoint PPT Presentation
1 / 98
Title:
From: Mathews Van Holde
Description:
Host of the electron transport chain (energy production) ... hydrophobicity scale: Based on partitioning in hexane. [conc. in phase 1] Kpartition ... – PowerPoint PPT presentation
Number of Views:242
Avg rating:3.0/5.0
Title: From: Mathews Van Holde
1
(No Transcript)
2
From Mathews Van Holde
3
Venter, J.C. et al (2001) The Sequence of the
Human Genome Science, (vol. 291, p. 1304
(2002)).
4
20-30 of all proteins are integral membrane
proteins
Some Functions of the Membrane Host of the
electron transport chain (energy
production). Semi-permeable barrier governing
cellular entry and exit. Host of most cellular
sensors. Platform for signal transduction. Shipp
ing containers. Site of cell-cell
junctions. Electrical device. Site of lipid and
oligosaccharide biosynthesis.
5
Nature Reviews Drug Discovery 1 727-730 (2002)
Marketed small-molecule drug targets by
biochemical class. GPCR, G-protein-coupled
receptor.
6
Explosion in the Numbers of Biomolecular
Structures Solved
Yet, only about 50 membrane protein structures
have been solved!
7
(No Transcript)
8
Membranes of a Gram Negative Bacterium
Gram positive bacteria and yeast do not have an
outer membrane, but do have a thick cell wall at
about the same location as the outer membrane.
from Alberts
9
(No Transcript)
10
(No Transcript)
11
Paintings of E. coli (left) and red blood
cell/blood by David Goodsell
12
The Singer-Nicholson Fluid Mosaic Model of
Biological Membranes Original formulation
(circa 1970) and recent updates.
13
U. of Texas
14
How Different Contributions to Molecular
Energetics Can Create Order Bilayer Formation in
Water
polar head group
apolar tail (acyl chains)
15
(No Transcript)
16
(No Transcript)
17
According to the original S-N Fluid Mosaic
Model Lipids and Membrane proteins are free to
undergo later diffusion in the plane of the
bilayer. However, flip flop across the
bilayer of proteins or lipids is very slow.
There is complete mixing in the bilayer plane of
all components.
18
Brownian motion and the random walk.
From Krell Institute
If you looked down onto a membrane surface and
watched the movements of a single lipid, this is
about what it would look like. Driven by kT
energy.
19
Membrane Proteins are Vectorially Distributed
with Respect to Membrane.
20
Lipids are also vectorially disposed
21
Asymmetric distribution of phospholipids between
the inner outer monolayers of erythrocyte
plasma membrane
22
Revisions to the original FMM Not all
membranous molecules undergo free lateral
diffusion.
23
(No Transcript)
24
Revision to FMM Sometimes, there are domains
which have a distinctive composition of
lipids and proteins. For example lipid rafts
are rich in cholesterol, sphingolipids, and
certain proteins. May have important functions.
25
(No Transcript)
26
Revisions to FMM Endocytosis is so extensive
that it means entire contents of plasma membrane
may be internalized within half an hour!
27
What are Lipids?
Simplest lipids are fatty Acids Myristic (C14,
saturated), palmitic (C16 saturated) and oleic
(C18, 1 double bond, unsaturated) acids (left to
right).
28
(No Transcript)
29
(No Transcript)
30
(No Transcript)
31
(No Transcript)
32
(No Transcript)
33
(No Transcript)
34
The glycosylation of the complement regulatory
protein, human erythrocytes CD59. P.M. Rudd,
B.P. Morgan, M.R. Wormald, D.J. Harvey, C.W. van
den Berg, S.J. Davis, M.A.J. Ferguson and R.A.
Dwek (1997) J. Biol. Chem., 272, 7229-7244.
blue N-linked gray O-linked yellow active site
residues green GPI anchor
35
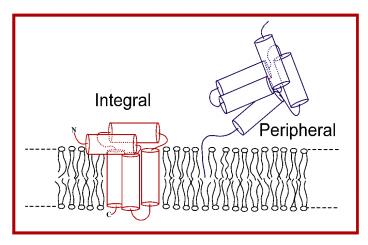
Varieties of Membrane Proteins Integral and
Peripheral
Not shown proteins bound to membrane surface via
electrostatic attraction between negatively
charged surface and positively charged protein
domains. (On the average about 1 out of 10 lipids
has a negatively charged head group.) Also not
shown proteins bound to membrane through an
amphipathic domain which dips below the membrane
surface, but which does not span the membrane.
36
(No Transcript)
37
Model Membranes Media in Which to Study Lipids
and Membrane Proteins
38
Multilamellar Vesicles Onion-Like. Each layer
one bilayer with a thin layer of water between
layers.
39
Unilamellar Vesicle AKA Liposome
40
Planar Lipid Membrane or Black Lipid
Membrane often used in electrophysiology.
41
Supported Monolayer and Bilayer. Block is solid
support to which lipid is adhered.
Monolayer at air/water interface. Tails in the
air.
42
Detergent Usually Conical Shape 2 short (6-8
carbons) Unsaturated acyl chains, or 1
alkyl/acyl Chain (8-14 carbons). Modest monomer
solubility in water. Micellar aggregates
have high solubility.
Lipid Cylinder Shape Usually 2
acyl/alkyl Chains, at least 12 carbons
each Extremely low monomer solubility.
43
How Different Contributions to Molecular
Energetics Can Create Order Bilayer Formation in
Water
polar head group
apolar tail (acyl chains)
44
Beta-octylglucoside
Dodecylsulfate (SDS)
Beta-dodecylmaltoside
CHAPS A bile salt-based detergent (Janus-like)
Triton X-100
45
(No Transcript)
46
Shapes of Micelles Most Micelles are NOT
Spherical
Prolate Ellipsoid
Oblate Ellipsoid
Bicelle
D. Small, The Physical Chemistry of Lipids
47
(No Transcript)
48
Detergents Vital Information
- Detergent Critical Micelle Concentration (CMC)
- When total detergent concentration is below
CMC, all detergent molecules - are monomeric in solution.
- Above CMC there is a free (monomeric) detergent
concentration equal to CMC - Above CMC there is a micellar detergent
concentration equal to - total detergent concentration CMC
- Aggregation Number the average number of
detergent molecules in a single - micelle.
- Concentration of micelles (total detergent
conc. CMC)/aggregation number
Aggregate Molecular Weight of Micelle
Aggregation number X detergent molecular
weight.
49
Temperature
From Calbiochem Info Brochure
50
(No Transcript)
51
(No Transcript)
52
(No Transcript)
53
(No Transcript)
54
The use of mild detergents for solubilizing,
purifying, and reconstituting functional membrane
protein systems. In this example, functional
Na-K pump molecules are purified and
incorporated into phospholipid vesicles. The
Na-K pump is an ion pump that is present in the
plasma membtane of most animal cells it uses the
energy of ATP hydrolysis to pump Na out of the
cell and K in, as discussed in chapter 11.
55
Bicelles Two Types 1 Based on
Bile Salt-Like Detergents 1 Based on Short Chain
PC
Structure 6, 1227-1234 (1998)
56
Lipid Bilayer Phases
57
A.K.A. Fluid Phase
- The "melting" of membrane lipids
- Below a certain transition temperature, Tm,
membrane lipids are rigid and tightly packed - Above the transition temperature, lipids are much
more flexible and mobile - The transition temperature is characteristic of
the lipids in the membrane
58
Gel Phases La Phase
59
Reithmeier Deber, pgs 337-393 in Deber
Phase Diagram for DPPC Bilayers
60
The melting temperature or phase transition
temperature, Tm, is reduced when When
saturated lipid tails are replaced by unsaturated
lipid tails. When shorter lipid tails replace
longer lipid tails. When cholesterol is
introduced. When ether-linked lipids are
replaced with ester-linked lipids. Native
membranes are almost always in the liquid
crystalline phase bilayer fluidity is required
for normal membrane function!
61
Bilayer Dimensions Lewis and Engelman, JMB 1983
DMPC Tm 24 deg. At 36 deg Phosphate to
Phosphate 3.4 nm Hydrophobic Thickness 2.3
nm Surface area 66 square angstroms DPPC
Tm 41 deg. At 44 deg Phosphate to phosphate
3.7 nm Hydrophobic Thickness 2.6
nm Surface area 67 square angstroms DOPC
Tm -14 deg. At 20 deg. Phosphate to
phosphate 3.8 nm Hydrophobic thickness
2.7 nm Surface area 70 square
angstroms EYPC (mostly POPC) Hydrophobic
thickness 2.8 nm E. coli lipids Phosphate to
phosphate ca. 4.2 nm
62
Davis, BBA 737 117-171 (1983)
Relative Positional Order of Carbons on C16
Acyl Chain of DPPC in Liquid Crystalline Phase
63
Weiner, Hristova, and White UC-Irvine
64
(No Transcript)
65
Role of Cholesterol Make membrane stronger
(less mobile than without) while helping to
prevent gel phase formation.
66
Why Membrane Proteins Are Membrane Proteins
67
conc. in phase 1 Kpartition
conc. in phase 2 DGopart
-RTlnKpartition
68
Goldman-Engelman-Steitz hydrophobicity
scale Based on partitioning in hexane.
conc. in phase 1 Kpartition conc. in
phase 2 DGopart -RTlnKpartition
Water-Octanol Partitioning gives idea of how
much side chain prefers to be just Under membrane
surface
Water-Hexane Partitioning gives idea of how much
side chain prefers to be in middle of bilayer
69
(No Transcript)
70
Rhodopsin
71
Helices of about 16-30 residues and beta barrels
can be stable in a membrane bilayer provided
that the side chains exposed to the hydrophobic
lipid phase are overwhelmingly hydrophobic.
72
Reithmeier Deber, pgs 337-393 in Deber
73
Hydropathy Plots
74
BR
75
Hydropathy Plots
76
Aromatic residues in membrane proteins are
usually found at the water-bilayer interface
rather than being deeply buried in the membrane.
Figure from P. van der Wels web
site http//comp.uark.edu/pvander/aromanch.html
77
The interiors of transmembrane domains of
membrane proteins (where protein-protein
interactions take place) are sometimes pretty
polar and sometimes pretty non-polar.
78
Apolar in both interior and exterior of barrel
79
OmpF apolar on exterior of barrel, but polar
interior
80
(No Transcript)
81
Sheets also have a sidedness residue side chains
alternate between the two sides of a sheet.
Outer membrane phospholipase M. Sansom, Oxford U.
82
FhuA Barrel is large enough to fit a whole other
protein domain inside. For some proteins of this
close domain may sometimes come out of the barrel
in a regulated manner.
83
(No Transcript)
84
(No Transcript)
85
Amphipathic helices have a polar face and a
non-polar face can sit on membrane surface.
S. White, U. Cal.-Irvine
86
(No Transcript)
87
Prostaglandin Synthase
88
Rhodopsin
89
GlpF
Glycerol Channel GlpF
90
(No Transcript)
91
(No Transcript)
92
ATPase Cartoon Here
93
(No Transcript)
94
(No Transcript)
95
Protein Folding In The Cell In vivo in a living
cell In vitro usually means cell has been
lysed, but many cellular components are present
still. Sometimes, this term is used for test
tube conditions involving purified molecules
leading to confusion.
From Mathews
96
The secretory pathway most integral membranes
are inserted into the membrane of the ER the
first stage of this pathway.
from Alberts
- all proteins which are secreted from the cell
(including components of extracellular matrix) - all membrane proteins of cell surface
- all soluble and membrane proteins destined for
ER, Golgi, endosomes, of lysosomes - the ER often takes up gt10 of the cellular volume
- probably 25-40 of all proteins either reside in
or pass through the secretory pathway
97
Hundreds of Hereditary Diseases (often Rare) are
Caused By Protein Misassembly in the Endoplasmic
Reticulum (ER) Examples of Disorders which stem
primarily from protein loss of function Cystic
fibrosis CF transmembrane regulator (a chloride
channel) Hereditary emphysema a-1 anti-trypsin
(a protease inhibitor) Tay-Sach b-hexosaminida
se (a degradative enzyme of lysosomes) Famil
ial hypercholesterolemia low density lipoprotein
(LDL) receptor (cell surface protein
involved in lipid/cholesterol clearance/metab
olism) retinitis pigmentosa rhodopsin and
other proteins of the photoreceptor
system Examples of Disorders which are
complicated by protein accumulation in the
ER Congenital hypothyroidism thyroglobulin
accumulates (goiter) Liver damage associated
with anti-trypsin accumulates in liver
cells emphysema osteogenesis imperfecta type I
pro-collagen hereditary hypofibrinogemia fibrino
gen (liver disease such as cirrhosis)
98
(No Transcript)